Mitose
Mitose is een type celdeling waarbij nieuwe dochtercellen ontstaan die gelijk zijn aan de moedercel, en op hun beurt in staat zijn om zich te delen. Tijdens dit celdelingsproces gaan de chromosomenparen in de celkern paarsgewijs uit elkaar, de kerndeling. Dit is een belangrijk onderdeel van de celcyclus.
 |
 |
 |
 |
In de kern van een eukaryote cel liggen de chromosomen met het erfelijke materiaal en de kernlichaampjes (nucleoli) ingebed in het kernplasma. De kern wordt omgeven door een dubbele membraan, de kernmembraan.
Een kern kan niet zomaar in tweeën worden gedeeld. De chromosomen moeten bij een mitose onveranderd worden doorgegeven van moedercel naar dochtercel. De chromosomen, die (vrijwel) alle erfelijke informatie van het organisme bevatten, worden dan ook verdubbeld of gekopieerd, en daarna over beide cellen verdeeld. Deze erfelijke informatie is belangrijk voor de regeling van (vrijwel) alle biologische activiteiten in de cel, en voor de ontwikkeling en de functie van de cel. De verdubbeling van de chromosomen vindt plaats tijdens de S-fase van de celcyclus, en maakt geen onderdeel uit van de mitose (de M-fase).
Een deel van de erfelijke informatie bevindt zich niet in de celkern, maar in de mitochondriën (het mitochondriaal DNA) en in de eventueel aanwezige plastiden (het chloroplast-DNA). Deze organellen ondergaan geen mitose, maar vermeerderen zich door binaire deling en knopvorming. Daarna verdelen ze zich over de dochtercellen.
Mitose werd in 1875 ontdekt toen biologen met een lichtmicroscoop cellen onderzochten.
De verschillende fasen
bewerkenProfase
bewerkenNa de interfase waarin het erfelijk materiaal verdubbeld is volgt de eerste fase van de mitose: de profase of voorbereidingsfase. De profase wordt gekenmerkt door:
- vorming van de spoelfiguur
- spiralisatie van en aanhechting van de chromosomen.
- verdwijnen van de nucleolus
In eukaryote cellen ligt net buiten de kernmembraan het spoellichaampje centrosoom, een gebied waarin bij dierlijke cellen ook de twee loodrecht op elkaar staande centriolen liggen. De centriolen en centrosomen verdubbelen zich in het begin van de profase en elk paar beweegt zich dan langzaam naar één kant van de kern. Geleidelijk ontwikkelt zich uit elk van de centrosomen een structuur van draden. Deze draden zijn gemaakt van microtubuli (Latijn: tubuli, buisjes). Een aantal evenwijdig lopende microtubuli vormt samen bundels, spoeldraden genaamd, die groot genoeg zijn om met de lichtmicroscoop te zien. De microtubuli zijn gemaakt van het eiwit tubuline. Er zijn drie soorten microtubuli: de astral (Engels), kinetochore en de interpolaire microtubuli. Als de centrosomen, bij de dierlijke cel ook met de centriolen, elk aan één kant van de kern aangekomen zijn, bij de 'polen' van de cel, is er een netwerk van draden ontstaan dat de gehele kern omspant. Dit wordt spoelfiguur genoemd. De spoelfiguur is voltooid aan het begin van de metafase. De functie van de centriolen is nog onbekend. Ook zonder centriolen kan een dierlijke cel zich delen.
Tegen het einde van de profase gaan de verdubbelde chromosomen, die nog kriskras verspreid en gekronkeld door de cel liggen, zich oprollen (spiraliseren). Door het verkorten van de chromosomen worden zij dikker en met een goede lichtmicroscoop is dan te zien dat elk chromosoom uit twee identieke delen bestaat, de chromatiden. De chromatiden zijn na het spiraliseren kort en dik genoeg om over de twee dochtercellen verdeeld te worden zonder dat ze breken. De chromatiden zitten aan elkaar vast in een gebied dat centromeer wordt genoemd. Elk van de chromatiden zit vast aan de draden van de spoelfiguur. Deze draden worden trekdraden genoemd. Door de spanning van de trekdraden aan elke kant van de chromatiden worden de chromosomen naar het midden van de cel gebracht. Een tweede soort draden van de spoelfiguur wordt steundraden genoemd, deze zitten niet vast aan de chromatiden, maar lopen door van centrosoom tot centrosoom.
-
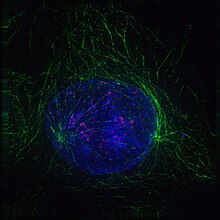 Cel in profase
Cel in profase -
 Cel in prometafase
Cel in prometafase -
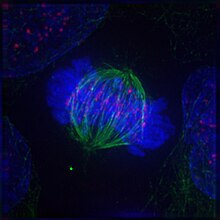 Cel in metafase
Cel in metafase



Prometafase
bewerkenTijdens de prometafase verbrokkelt de kernmembraan. De microtubuli van het spoellichaampje kunnen nu doordringen in de kern en interactie aangaan met de chromosomen. De chromosomen zijn in de prometafase volledig gespiraliseerd. Op beide chromatiden van elk chromosoom ontwikkelt zich bij het centromeer (een insnoering in het chromosoom) een structuur van eiwitten (het kinetochoor). Sommige microtubuli van de spoelfiguur hechten zich aan de kinetochoren: de kinetochoormicrotubuli. Dit gaat gepaard met schokkerige bewegingen van de chromosomen. De andere microtubuli werken in op de tegenovergelegen microtubuli van de spoelfiguur, waardoor de cel langer wordt van vorm.
Metafase
bewerkenTijdens de metafase zijn de centrosomen, die het spoelfiguur vormen, volledig naar de uiteinden (polen) van de cel bewogen. De centromeren van elk van de chromosomen hebben zich in één vlak in het centrum van de cel geordend, in het zogenoemde equatorvlak (metaphase plate). De kinetochoormicrotubuli hechten zich aan de kinetochoren van elk chromatide. De metafase is de fase waarin de chromosomen het duidelijkst bestudeerd kunnen worden, deze chromosomen heten metafase-chromosomen.
Anafase
bewerkenDe beslissende fase van de mitose, die slechts enkele minuten duurt, is de anafase. De verbindingen tussen de chromatiden (bij de centromeren) laten los en de zusterchromatiden (nu chromosomen) bewegen elk in tegengestelde richting naar de polen doordat de spoeldraden ze naar de polen trekken. Dit gebeurt doordat de trekdraden verkorten. Doordat de microtubuli gehecht zijn aan de centromeren, worden de centromeren vóór de armen uit getrokken, zodat de chromosomen V-vormig zijn in deze fase. De anafase stopt wanneer elke set chromosomen, die met een snelheid van ongeveer 1 μm/min bewegen, aangekomen is aan zijn pool. Hier zullen de nieuwe kernen gevormd worden. Naar welke pool de chromatiden zich begeven, is willekeurig.
-
 Cel in anafase
Cel in anafase -
 Longcel van een watersalamander in anafase
Longcel van een watersalamander in anafase -
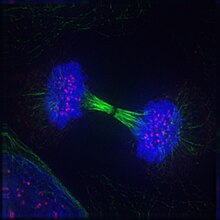 Cel in telofase
Cel in telofase



Telofase
bewerkenTijdens de telofase despiraliseren de chromatiden en worden weer lange dunne draden (chromatine), die niet van elkaar te onderscheiden zijn met de lichtmicroscoop. De nucleoli verschijnen weer. De chromatiden worden opnieuw 'verpakt' in een kernenvelop en de vorming van twee complete kernen is dan voltooid. Alle resterende microtubuli van de spoelfiguur depolymeriseren. Nadat de telofase ten einde is gekomen, is het resultaat twee nieuwe kernen. Hiermee is de kerndeling beëindigd.
Cytokinese
bewerkenHet cytoplasma en de organellen worden tijdens de cytokinese evenredig verdeeld over de twee dochtercellen. Cytokinese overlapt met de telofase, waardoor de cellen na insnoering elkaar loslaten en er meteen nieuwe aparte cellen ontstaan, ieder met een eigen celmembraan.
Duur van de verschillende fasen
bewerkenDe duur van de verschillende fasen is afhankelijk van het celtype en het organisme. Gemiddeld duurt in een celcultuur een celdeling bij cellen van zoogdieren 12 tot 24 uur.
Voorbeelden van verschillende organismen (duur in uren):
| Celtype | G1 | S-fase | G2 | Mitose | Totale duur van de celdeling in uren |
|---|---|---|---|---|---|
| Slijmzwam (Physarum polycephalum) | zeer kort | 2 | 4 | 0,7 | ca. 6,7 |
| Tuinboon (Vicia faba) Meristeem van het wortelpuntje | 4 | 9 | 3,5 | 2 | 18,5 |
| Muis (Mus musculus) Tumorcellen in een cultuur | 10 | 9 | 4 | 1 | 24 |
| Mens (Homo sapiens) Tumorcellen in een cultuur | 8 | 6 | 4,5 | 1 | 19,5 |
Factoren die de mitose beïnvloeden
bewerkenColchicine is een mitose-remstof, een antimitoticum, die bij de celdeling de overgang van de metafase naar de anafase verhindert, waardoor de vorming van het spoelfiguur geblokkeerd wordt en de chromosomen niet uit elkaar getrokken kunnen worden. Door behandeling met colchicine werden vooral in het verleden tetraploïde planten ontwikkeld. Bijvoorbeeld tetraploïd Engels raaigras (Lolium perenne), dat gebruikt wordt voor de inzaai van grasland. Andere antimitotica zijn onder meer Softenon en Taxol.
Mitotica, of mitogenen, zijn stoffen die de mitose bevorderen. Voorbeelden hiervan zijn het groeihormoon in de hypofyse en het schildklierhormoon, thyroxine. Dit zijn chemische factoren die de mitose beïnvloeden. Uiteraard zijn er dan ook fysische factoren. Deze zijn licht (de mitose bij planten is seizoensgebonden), temperatuur (minimum – optimum – maximum) en straling ( röntgen- en gammastralen kunnen de delende cel doden, of misvormen, wat kan leiden tot kanker).
Ascomyceten
bewerkenAscomyceten hebben een complexe vorm van mitose. Bij veel soorten bestaan in het geheel geen seksuele stadia of zijn deze niet bekend. Wel komt er paraseksualiteit voor. Paraseksualiteit houdt een fenomeen in waarbij het zonder eigenlijke seksuele gebeurtenissen tot een versmelting van twee celkernen en daardoor een verdubbeling van het aantal chromosomen komt. Daarna vindt een complexe vorm van mitose plaats, waarbij het tot een crossing-over (overkruising), een uitwisseling van genetisch materiaal tussen twee op elkaar passende chromosomen komt. Bij vormen van geslachtelijke voortplanting treedt overkruising daarentegen alleen bij de meiose op. Uiteindelijk ontstaan door haploïdisatie weer twee celkernen met hun eigen chromosomenreeks, die zich nu echter genetisch van de beide uitgangskernen onderscheiden.