Geslachtelijke voortplanting bij Embryophyta
De geslachtelijke voortplanting bij de Embryophyta (of landplanten) is de voortplanting waarbij twee ouder-organismen, een vrouwelijke en een mannelijke, betrokken zijn. Deze voortplanting verschilt nogal per groep.
Bij mossen, levermossen, hauwmossen en varens is deze voortplanting veel eenvoudiger dan bij naaktzadigen en bij bedektzadigen. Het proces van geslachtelijke voortplanting kan goed begrepen worden door een onderlinge vergelijking van de levenscyclus en het voortplantingsproces bij mossen, varens, naakt- en bedektzadigen.
Generatiewisseling en kernfasewisseling bewerken
Een belangrijk begrip is generatiewisseling, zoals in 1849 voor het eerst beschreven door Wilhelm Hofmeister.
Anders dan de verwante algengroepen hebben alle Embryophyta (landplanten) een biologische levenscyclus met een generatiewisseling en een kernfasewisseling: twee meercellige generaties wisselen elkaar af: een diploïde generatie wisselt af met een haploïde generatie. De generaties worden genoemd naar de voortplantingscellen die zij produceren.
De haploïde gametofyt produceert de gameten (geslachtelijke voortplantingscellen) door mitose (gewone celdeling). Na versmelting van de haploïde gameten ontstaat een diploïde zygote die uitgroeit tot sporofyt.
De diploïde sporofyt produceert door meiose (reductiedeling, rijpingsdeling) de sporen (of meiosporen).
Evolutionaire trends bewerken
Bij de geslachtelijke voortplanting Embryophyta zijn verschillende evolutionaire trends te herkennen, met name op het gebied van de afwisseling van individuen in hun levenscyclus, de mate van ontwikkeling van de sporofyten, de sporangia en de sporen, de wijze van bevruchting en de voortplanting.
Afwisseling van individuën bewerken
Bij mossen, levermossen en hauwmossen is er in de levenscyclus slechts één individu - het zijn daarom haplobionten. De "mosplant" is de dominante haploïde gametofyt, waarop de diploïde sporofyt groeit.
Varens hebben meestal een duidelijke afwisseling van individuen: het zijn per definitie diplobionten. De zelfstandig levende diploïde sporofyt is dominant (het best ontwikkeld en met de langste levensduur). Er is een kleinere, vrij levende, haploïde gametofyt. De gametofyt ontwikkelt zich in sommige gevallen binnen de tegen uitdroging beschermende sporewand.
Zaadplanten, zoals de naakt- en bedektzadigen met steeds slechts één individu, zijn haplobionten, maar nu is de "plant" de sporofyt. De (mannelijke) microgametofyt ontwikkelt zich binnen de sporewand tot een stuifmeelkorrel. Het stuifmeel wordt door de wind of door andere agentia verspreid, zodat deze terechtkomen bij de vrouwelijke voortplantingsorganen, waar ze kiemen en zorgen voor de bevruchting. De (vrouwelijke) macrogametofyt blijft volledig binnen het diploïde macrosporangium (nucellus) binnen de moederplant.
Sporofyten bewerken
Als gekeken wordt naar de sporofyten, ziet men bij de mossen een onvertakte sporofyt met één eindstandig sporangium (monosporangiaat), dat veel gelijkvormige sporen vormt. Daarentegen is bij varens de sporofyt vertak en draagt meerdere sporangia (polysporangiaat). Bij sommige varens is er een differentiatie in mannelijke micro- en vrouwelijke macrosporangia, waar respectievelijk de microsporen en de macrosporen worden gevormd. Sporofyten zijn veel meer dan gametofyten bestand tegen uitdroging door het bezit van een cuticula op de epidermis en huidmondjes die de vochthuishouding regelen.
Bevruchting bewerken
Ook bij de bevruchting zelf is een evolutionaire tendens waarneembaar. Bij mossen, varens en verschillende groepen naaktzadigen zijn de mannelijke voortplantingscellen, de antherozoïden, voorzien van flagellen, soms van cilia. De beweeglijke zaadcellen bevruchten de onbeweeglijke eicel. Hier spreekt men van oögamie. Bij andere groepen naaktzadigen en bij bedektzadigen groeit de pollenkorrel uit als een pollenbuis en brengt zo de mannelijke celkernen bij de eicel. Hier spreekt men van sifonogamie.
Disseminatie: sporen en zaden bewerken
De voortplanting bij mossen en varens vindt plaats door de in grote aantallen gevormde sporen. Bij mossen groeit uit een enkele spore een voorkiem, waarop een groot aantal mosplantjes gevormd kunnen worden, die elk weer een of enkele sporofyten (sporenkapsels) vormen. Bij varens worden de sporen gevormd in sporangia die in grote aantallen worden gevormd op een varenplant. Uit een enkele spore groeit een voorkiem, en daarop een of enkele sporofyten (varenplanten).
Bij de zaadplanten (de naakt- en bedektzadigen) blijven de macrosporen van de moederplant binnen het macrosporangium (of nucellus), dat omhuld wordt door integumenten. Het macrosporangium met integumenten samen vormen een zaadknop. De zaadknoppen zijn zeer talrijk aanwezig op de moederplanten. De macrosporen ontwikkelen zich ter plaatse tot een macrogametofyt (of kiemzak). Na de bevruchting ontwikkelt zich het kiempje (embryo), dat ook beschut blijft binnen de macrogametofyt. Bij rijpheid valt het geheel af: de tot zaden gerijpte zaadknoppen, bij bedektzadigen al of niet samen met omhullende structuren, die dan vrucht worden genoemd. Nu kan het kiempje zich verder ontwikkelen en uitgroeien tot een nieuwe plant.
Hieronder wordt, na een algemene behandeling, de geslachtelijke voortplanting kort besproken per verwantschapsgroep: de mossen met verwanten, de varens met verwanten, de verschillende groepen naaktzadigen en de bedektzadigen.
Leven op het land en levenscyclus bewerken
| Stamboom | ||
|---|---|---|
|
Leven op het land bewerken
De Embryophyta hebben als groep het land veroverd en zijn daar de meest dominante groep van organismen: ze zorgen voor verreweg het grootste aandeel in de primaire productie van de biomassa. De Embryophyta kunnen de droogte op het land beter overleven, en ze hebben een efficiëntere fotosynthese dan de zustergroepen die behoren tot de kranswieren en de groenwieren.
De Embryophyta hebben in de evolutie hun afhankelijkheid van water bij de voortplanting overwonnen. De onafhankelijkheid van het water begint met de evolutie van planten met eencellige sporen met een door sporopollenine waterafsluitende celwand. Planten hebben gedurende de sporofytfase een epidermis met een waterafsluitende cuticula en ze kunnen met huidmondjes de waterafvoer en de gaswisseling van zuurstof (O2) en van kooldioxide (CO2) regelen. Ook doordat veel soorten een symbiotische relatie met een schimmel aangegaan in de vorm van een mycorrhiza, kunnen deze beter in hun waterbehoefte voorzien. Dergelijke mycorrhiza's vormen uitgebreide ondergrondse netwerken van hyfen (schimmeldraden), met behulp waarvan de planten water met de daarin opgeloste mineralen kunnen opnemen.
Hoewel de groep als geheel "landplanten" wordt genoemd, zijn er veel soorten die secundair zijn aangepast aan het leven in het zoete, brakke of zelfs het zoute water, zoals mangrovesoorten en zeegrassen.
Levenscyclus bewerken
Wilhelm Hofmeister onderkende omstreeks 1849 als eerste de overeenkomst in de generatiewisseling van mossen (en verwanten), varens (en verwanten) en zaadplanten, met name de coniferen, en de homologie van de organen voor de voortplanting bij verschillende groepen van de Embryophyta (landplanten).
De Embryophyta hebben alle een levenscyclus met een afwisseling van twee generaties en met een afwisseling van kernfases: met een haploïde generatie, de gametofyt (de gametenvormende generatie) wisselt af met een diploïde generatie, de sporofyt (de sporenvormende generatie).[1] Een levenscyclus met een dergelijke afwisseling van generaties heet een digenetische cyclus.
Bij de Embryophyta begint de haplofase (haploïde kernfase) of gametofyt-fase met de door meiose (reductiedeling, rijpingsdeling) gevormde eencellige sporen, die uitgroeien tot gametofyt (de generatie die de haploïde gameten of geslachtscellen vormt). De diplofase (diploïde kernfase) of sporofytfase begint met de diploïde zygote (de bevruchte eicel), die zich via het embryo-stadium ontwikkelt tot een meercellige sporofyt (de generatie die de sporen vormt).
| De belangrijkste processen in de levenscyclus zijn de bevruchting en de (sporische) meiose. Hiertussen vinden de groei en ontwikkeling plaats tot een meercellig individuen: de gametofyt en de sporofyt die elkaar in een cyclus afwisselen, de digenetische cyclus.[2][3][4][5] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
In het algemeen zijn er in de levenscyclus van Embryophyta de volgende processen en de stadia van groei en ontwikkeling te onderscheiden:
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Het bovenstaande laat het principe van generatiewisseling met kernfasewisseling zien: de gametofyt wisselt af met de sporofyt (de sporenvormende generatie); dit type generatiewisseling is de digenetische cyclus. De haploïde generatie (haplofase) wisselt hier af met een diploïde generatie (diplofase); een organisme met een dergelijke kernfasewisseling is een diplohaplont.
De meiose of reductiedeling vindt plaats bij de vorming van de sporen (en niet bij de vorming van de gameten); dit heet sporische meiose. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Verscheidenheid in levenscycli bewerken
Er bestaat bij Embryophyta een grote verscheidenheid in de onderlinge verhoudingen tussen sporofyt en gametofyt. Bij mossen, levermossen en hauwmossen is de gametofyt de dominante generatie, bij de overige Embryophyta is de sporofyt dominant. Dominant wil hier zeggen: beter ontwikkeld, met langere levensduur en in de groei en ontwikkeling onafhankelijk van de andere generatie. Soms is daarbij sprake van afwisseling van individuen, als zowel de sporofyt als de gametofyt onafhankelijk van elkaar zelfstandig kunnen leven, zoals bij veel varens.
Bevruchting en zygote-vorming bewerken
|
mossen s.l., varens, paardestaarten | |||||||||||
|
palmvarens | |||||||||||
|
eenhuizige tweeslachtige zaadplanten | |||||||||||
|
tweehuizige zaadplanten | |||||||||||
| ||||||||||||
De meest oorspronkelijke landplanten en hun huidige nakomelingen, zoals de mossen, zijn voor de oögamie afhankelijk van water: de mannelijke gameet, de antherozoïde, zaadcel of spermatozoïde heeft twee flagellen en zwemt in een aanwezige waterfilm naar het archegonium en bevrucht de daarin aanwezige, onbeweeglijke vrouwelijke gameet, de eicel.
Bij heterospore varens ligt de eicel in een archegonium, dat gevormd wordt in het binnen de wand van de macrospore gegroeide macroprothallium. Ook het microprothallium met de antheridia ontwikkelt zich al binnen de wand van de microspore, maar de antherozoïden moeten nog naar het archegonium zwemmen.
Bij de palmvarens, waarvan de zeer kleine microsporen in de pollenkamer van de nucellus (macrosporangium) kiemen, moeten de antherozoïden nog maar een zeer kleine afstand zwemmen naar het archegonium.
Bij sommige naaktzadigen en de bedektzadigen kunnen de mannelijke gameten niet zwemmen en is de bevruchting geheel onafhankelijk van water. In de rudimentaire antheridia worden de mannelijke gameten gevormd zonder flagellen. Ze bestaan uit zaadcelkernen die door een pollenbuis naar de eicel worden gebracht; dit heet sifonogamie .
De bestuiving en bevruchting vinden hier plaats door wind- of insectenbestuiving, gecombineerd met sifonogamie en zijn zo onafhankelijk geworden van de aanwezigheid van water.
In sommige gevallen, bijvoorbeeld bij de palmvarens en tweehuizige zaadplanten kan men spreken van mannelijke en vrouwelijke zygoten, als deze zich ontwikkelen tot mannelijke of vrouwelijke planten. Het verschil in geslacht wordt geregeld door geslachtschromosomen.
Sporofyten en sporangia bewerken
|
mossen s.l. | ||||||||||||||||||
|
wolfsklauwen Lycopodium | ||||||||||||||||||
|
paardestaarten Equisetum | ||||||||||||||||||
|
isospore varens | ||||||||||||||||||
|
Selaginella, Marsilea, heterospore varens | ||||||||||||||||||
|
tweeslachtige eenhuizige zaadplanten | ||||||||||||||||||
|
tweehuizige zaadplanten | ||||||||||||||||||
| |||||||||||||||||||
Bij Embryophyta ontwikkelt de diploïde zygote (2n), het product van de bevruchting van de eicel door een antherozoïde (of door een mannelijke kern), zich door tot een door de macrogametofyt omgeven meercellig embryo. Het embryo groeit verder tot een meercellige sporofyt. Sporofyten zijn duidelijk beter dan de gametofyten beschermd tegen uitdroging door de aanwezigheid van een epidermis (een één cellaag dikke opperhuid) met een cuticula, bestaande uit wasachtige bestanddelen.
Bij de eenvoudigste landplanten, zoals bij de mossen en levermossen, leeft de sporofyt op de gametofyt (de "mosplant"). De sporofyt bestaat uit een voet en een solitair sporenkapsel (sporangium of sporogoon), gedragen aan het einde van een onvertakte (en onbebladerde, maar soms wel fotosynthetische) kapselsteel of seta. De sporofyt heeft soms eigen mogelijkheden tot fotosynthese (chloroplasten) en gaswisseling (huidmondjes), maar gewoonlijk worden energie en voeding van de gametofyt opgenomen via de voet. De mosplanten nemen hun energie en voeding (licht, CO2, water en mineralen) op met het gehele oppervlak van de blaadjes, stengels en rizoïden.
Bij de verder geëvolueerde landplanten, behorende tot de clade van de "polysporangiaten" zoals bij varens en zaadplanten, staan de sporangia gegroepeerd. De sporofyt is bij varens vertakt en vormt een vrij snel zelfstandige "varenplant". Hun energie en voeding worden door planten bovengronds met behulp van bladeren (licht en CO2) en ondergronds door wortels (water en mineralen) opgenomen. De "varenplant" draagt de sporangia vaak in grote aantallen in sporangiënhoopjes (sori) op de sporofyllen of sporangioforen.
Bij zaadvarens zijn macrosporangia met cupula te vinden aan grote, fertiele bladen. Een cupula is een kraag van onder het macrosporangium staande, minder of meer vergroeide zijassen. Een cupula omgeeft gedeeltelijk tot grotendeels een of enkele macrosporangia.
Bij zaadplanten omgeven een of twee integumenten het macrosporangium, dat hier nucellus wordt genoemd. De integumenten zijn enigszins te vergelijken met een cupula, maar het is niet duidelijk of het om homologe structuren gaat. Een nucellus met integumenten heet zaadknop (ovulum). De zaadknoppen staan op macrosporofyllen in de oksel van een zaadschub (een schubvormig schutblad) of erbovenop. De integumenten laten aan de top een opening vrij waardoor microsporen of pollenkorrels naar binnen kunnen. De voortplanting wordt bij zaadplanten verzorgd door de grote aantallen zaadknoppen (die elk een nucellus bevatten), die bij bevruchting de zaden kunnen vormen. Hier spreekt men van geslachtelijke voortplanting door middel van zaden. De zaadknoppen worden omgeven door een vruchtbeginsel, dat bestaat uit één of enkele carpellen (vruchtbladen).
Sporangia, meiose en sporevorming bewerken
|
monoecische mossen, isospore varens | ||||||||||||||
|
dioecische mossen, paardestaarten | ||||||||||||||
|
Selaginella, Marsilea, heterospore varens | ||||||||||||||
|
naaktzadigen | ||||||||||||||
|
bedektzadigen | ||||||||||||||
| |||||||||||||||
Sporen zijn losse, dikwandige cellen zonder eigen mogelijkheden zich voort te bewegen. De celwand van de sporen is stevig en ondoorlatend voor water door de aanwezigheid van sporopollenine, zodat ze uitdroging beter overleven. Evenals geslachtscellen (gameten) zijn sporen haploïde, maar sporen kunnen uitgroeien tot een meercellige generatie, de gametofyt.
Sporen worden door reductiedeling of meiose gevormd in meercellige sporangia uit sporemoedercellen (sporocyten). Hierbij wordt uit de diploïde moedercel (met 2n chromosomen) vier haploïde dochtercellen gevormd (met 1n chromosomen). Hierom worden deze sporen ook wel meiosporen genoemd, in tegenstelling tot de mitosporen die door mitose worden gevormd.
Bij de isospore (dat wil zeggen: met sporen van gelijke grootte) landplanten met de meer oorspronkelijke kenmerken (zoals bij mossen s.l. en bij veel varens) worden de sporen in grote aantallen gevormd. De zeer talrijke sporen zijn klein en worden door de wind verspreid. De sporen hebben een belangrijke rol bij de ongeslachtelijke voortplanting.
Vrijwel alle mossen s.l. (dat wil zeggen inclusief de levermossen en hauwmossen) en de meeste varens zijn isospoor: de haploïde eencellige sporen die alle van gelijke grootte zijn worden gevormd in isosporangiate sporangia (er is maar één type sporangium). De sporen worden gevormd door meiose , meestal in tetraden (viertallen). Uit de sporen groeien bij varens de prothallia (voorkiem), die antheridia en archegonia vormen, maar bij mossen kiemen de sporen en groeien eerst uit tot een protonema. Op de protonemata groeien de een- of tweeslachtige mosplanten, die antheridia en/of archegonia vormen. Voor bevruchting is water nodig zodat de antherozoïden naar de eicel kunnen zwemmen.
Bij veel soorten is er slechts een type sporangia (isosporangiaat) en zijn de sporen van gelijke grootte (isosporie). In de microsporangia en de macrosporangia van enkele groepen van varens en bij de zaadplanten worden de kleine, mannelijke microsporen en de grotere, respectievelijk vrouwelijke macrosporen gevormd, zodat er ook twee typen van sporen te onderscheiden zijn (heterosporie).
Bij de paardenstaarten komt "functionele heterosporie" voor: ze zijn morfologisch isospoor maar fysiologisch heterospoor. De helft van de sporen (de androsporen) vormen mannelijke prothallia met antheridia en de andere helft van de sporen (de gynosporen) vormen de prothallia met archegonia.
Microsporangia en macrosporangia komen onder andere voor bij de Salviniales (een orde van de varens) en bij de zaadplanten.
Alle verdere plantengroepen zijn heterospoor en heterosporangiaat, dat wil zeggen: in microsporangia worden uit microsporemoedercellen (microsporocyten) door meiose de kleine, mannelijke sporen, microsporen gevormd, en in macrosporangia worden uit macrosporemoedercellen (macrosporocyten) de grote, vrouwelijke macrosporen gevormd. Deze varens zijn dus ook heterosporangiaat.
Met het ontstaan van de geslachtsverdeling met microsporen en macrosporen is de rol bij de ongeslachtelijke voortplanting in de loop van de evolutiegeschiedenis steeds verder verloren gegaan.
In de helmknoppen, een onderdeel van de meeldraden van de bedektzadigen, kunnen grote aantallen microsporen gevormd worden (het stuifmeel), maar in het macrosporangium, dat een onderdeel vormt van de zaadknop, wordt slechts nog een functionele macrospore gevormd. Bij bedektzadigen zijn de microsporangia dan ook aanzienlijk groter dan de macrosporangia.
De microsporen worden veelal door de wind verspreid. De microsporen kiemen en groeien uit tot een microprothallium met enkele antheridia. De kieming van de macrosporen vindt soms plaats in het macrosporangium waar ze gevormd zijn en binnen de celwand van de macrosporen. Daarbij worden ook archegonia gevormd, elk met een eicel. Voor bevruchting is gewoonlijk water nodig zodat de antherozoïden naar de eicel kunnen zwemmen.
De macrosporangia vormen slechts weinig macrosporen; soms is er slechts een macrosporemoedercel die zich meiotisch deelt waarbij een tetrade van macrosporen wordt gevormd. Van de vier, meestal op een rij liggende macrosporen gaan er drie te gronde en slechts een ontwikkelt zich verder tot een macroprothallium.
Verder geëvolueerde landplanten zijn heterospoor, met in grote aantallen in microsporangia geproduceerde (kleinere, mannelijke) microsporen en in kleine aantallen per macrosporangium geproduceerde (grotere, vrouwelijke) macrosporen. De sporen kiemen al binnen het sporangium en binnen de sporewand.
Bij de bedektzadigen wordt per zaadknop in het macrosporangium (nucellus) gewoonlijk slechts één functionele macrospore gevormd, die uitgroeit tot een uit enkele celkernen bestaande macrogametofyt.
Gametofyten, gametangia en gameten bewerken
|
mossen s.l. (inclusief levermossen & hauwmossen) | |||||||||||||||||||
| ||||||||||||||||||||
|
isospore varens | |||||||||||||||||||
|
paardestaarten | |||||||||||||||||||
|
heterospore varens, palmvarens | |||||||||||||||||||
|
coniferen, bedektzadigen | |||||||||||||||||||
| ♂ = mannelijk, ♀ = vrouwelijk, ⚥ = tweeslachtig | ||||||||||||||||||||
Uit de haploïde sporen groeien de haploïde gametofyten waarop de gametangia (voortplantingsorganen) staan: de mannelijke antheridia en vrouwelijke archegonia.
Bij veel mossen, levermossen en hauwmossen is er geen onderscheid tussen mannelijke en vrouwelijke sporen en mosplanten, maar er zijn veel soorten die wel een dergelijke geslachtsverdeling hebben: de gametofyten zijn tweehuizig. Een bekend voorbeeld daarvan is parapluutjesmos (Marchantia polymorpha). Mossen, levermossen en hauwmossen en veel varens zijn isospoor: er is geen onderscheid in grote vrouwelijke en kleine mannelijke sporen, hoewel er bij sommige tweehuizige mossen wel heterosporie bestaat.
Bij mossen groeien sporen uit tot een vaak draadvormige voorkiem, het protonema. De voorkiem is soms echter meer een celklompje. Hierop ontwikkelen zich de mosplantjes, die de gametangia vormen. Bij isospore varens is de voorkiem thalleus, vaak een groen vliezig blaadje, waarop de antheridia en archegonia worden gevormd.
Bij heterospore planten, zoals bij sommige varens en alle zaadplanten, vormen zich vrouwelijke macrosporen en mannelijke microsporen. Deze kiemen gewoonlijk binnen de sporewanden, de microgametofyten, respectievelijk de macrogametofyten.[7]
Gametangia hebben bij Embryophyta een buitenste cellaag van steriele cellen. De in de gametangia gevormde gameten worden beschermd door een buitenste, eromheen liggende, steriele laag cellen. De antheridia (mannelijke gametangia), die de antherozoïden vormen, en de archegonia (vrouwelijke gametangia), die elk een eicel bevatten, zijn in de meest oorspronkelijke situatie (zoals bij mossen en veel varens) in grotere aantallen vrijstaand op de zelfstandige en vrijlevende gametofyt. Bij verder geëvolueerde landplanten is een tendens van steeds verdere reductie van de gametangia. De prothallia vormen slechts 1 of enkele archegonia per prothallium, met een zeer korte archegoniumhals, die soms zelfs afwezig is. Er is dan nog één ventrale halskanaalcel. Bij zaadplanten bestaan de gametangia uit enkele cellen of celkernen. De antheridia en archegonia zijn daar microscopisch klein en nog met moeite te herkennen. De antheridia en vooral de archegonia worden in de evolutionaire geschiedenis met hun gametangia en gametofyten opgenomen in omhullende beschermende structuren.
De gameten (geslachtscellen, geslachtelijke voortplantingscellen) worden gevormd in veelcellige gametangia (geslachtsorganen) met een buitenwand van steriele cellen. Het antheridium (mannelijke gametangium) produceert veel mannelijke gameten, de spermatozoïden of antherozoïden, die om te zwemmen ten tijde van de bevruchting water nodig hebben. Het archegonium (vrouwelijke gametangium) bevat slechts één onbeweeglijke vrouwelijke gameet, de eicel.
Geslachtsverdeling bewerken
| Geslachtsverdeling bij landplanten (Embryophyta) | |||
|---|---|---|---|
| Sporofyt en sporangia ↓ |
Gametofyt, verdeling gametangia | ||
| ⚥ Monoecisch (eenhuizig), Tweeslachtig |
♂ of ♀, Dioecisch (tweehuizig), Eenslachtig | ||
| Iso- sporangiaat { |
Isospoor → | eenhuizige mossen[8] wolfsklauwen isospore varens |
tweehuizige mossen |
| Heterospoor → | Ø | paardestaarten heterospore mossen | |
| Hetero- sporangiaat { en heterospoor |
Eenhuizig, → ⚥, Tweeslachtig |
heterospore varens eenhuizige zaadplanten | |
| ♂ of ♀, → Tweehuizig |
tweehuizige zaadplanten | ||
Met de term "geslachtsverdeling" kunnen twee verschillende verschijnselen worden bedoeld:
- de plaatsing van de mannelijke en vrouwelijke gametangia (de antheridia en archegonia) op de gametofyten
- de plaatsing van de mannelijke en vrouwelijke sporangia of sporendoosjes (micro- en macrosporangia) op de sporofyten.
Op grond van de verscheidenheid van grootte van sporangia maakt men onderscheid in isosporangiate en heterosporangiate planten:
Isosporangiate planten hebben sporangia, die alle van gelijke vorm en grootte zijn. Dit is het geval bij de meeste mossen en bij veel varens. Isosporangiate planten zijn vrijwel alle ook isospoor, waarbij de sporen uitgroeien tot tweeslachtige gametofyten die zowel antheridia als archegonia dragen, of, zoals bij sommige groepen van mossen, ze groeien uit tot mannelijke gametofyten en vrouwelijke gametofyten (tweehuizige mossen).
Bij bepaalde isosporangiate varens, zoals bij paardenstaarten, zijn de sporen morfologisch isospoor, maar functioneel heterospoor: uit de helft van de sporen groeien mannelijke planten, uit de andere helft groeien vrouwelijke planten.
Sommige isosporangiate mossen zijn morfologisch min of meer heterospoor, waarbij de kleinere sporen mannelijke mosplanten opleveren (die soms dwerggroei vertonen), en de grotere sporen vormen de vrouwelijke mosplanten.
Heterosporangiate planten zijn ook heterospoor: ze hebben microsporangia waar de microsporen gevormd worden en macrosporangia waar de macrosporen gevormd worden. Eenhuizige planten, waar de vrouwelijke macrosporangia of zaadknoppen en de mannelijke microsporangia of meeldraden op dezelfde plant gevormd worden, heten tweeslachtig. Er zijn ook eenslachtige soorten, waar de macrosporangia of zaadknoppen en de microsporangia of meeldraden op verschillende planten worden gevormd; er is dan sprake van mannelijke en vrouwelijke planten.
Mossen en varens bewerken
Bij mossen en varens is er een duidelijke overeenkomst in voortplantingsorganen, hoewel hun levenscyclus sterk lijkt te verschillen. Bij varens is de variatie groot. Het eenvoudiger proces van de voortplanting bij mossen en varens wordt eerst beschreven om daarna de voortplanting bij zaadplanten te verduidelijken.
Mossen, levermossen en hauwmossen bewerken
|
| |
|
Mossen, levermossen en hauwmossen hebben een heteromorfe, digenetische cyclus met dominante gametofyt: van de twee ongelijk gebouwde generaties is de haploïde gametofyt dominant (de mosplant, gametofoor), en leeft de sporofyt op, en afhankelijk van de gametofyt. De sporofyt bestaat met de voet in de mosplant bevestigde de kapselsteel of seta met aan de top deaarvan het sporenkapsel of sporogoon.
Door meiose worden in het sporogoon (sporenkapsel) door sporemoedercellen (sporocyten) de sporen gevormd. Via een protonema-stadium groeien de sporen uit tot een gametofyt, die bij deze groepen de dominante generatie is. De "mosplant" is dan ook de haploïde gametofyt. Mossen, levermossen en hauwmossen zijn over het algemeen isospoor: dat wil zeggen dat alle sporen van gelijke grootte zijn, maar deze kunnen desondanks bij sommige soorten uitgroeien tot gescheiden mannelijke en vrouwelijke mosplanten.
Heterosporie komt slechts bij enkele groepen van mossen voor. De mannelijke microsporen zijn kleiner en de vrouwelijke macrosporen zijn groter. Uit de mannelijke sporen groeien uiteindelijk de mannelijke mosplanten, die soms aanzienlijk kleiner zijn ("dwergmannetjes") dan de vrouwelijke mosplanten. Soms echter zijn de mannelijke planten beter ontwikkeld.[13]
De mannelijke geslachtsorganen (gametangia) zijn de antheridia en de vrouwelijke geslachtsorganen zijn de archegonia. De antheridia en archegonia hebben een beschermende wand van steriele cellen. Ze staan bij eenhuizige planten op dezelfde plant, maar bij tweehuizige planten staan deze op mannelijke of op vrouwelijk planten. Heterosporie komt echter bij mossen zelden voor.
Het antheridium is een in het thallus kort gesteeld, zittend of verzonken, bol- tot eivormig orgaantje met een wand van steriele cellen. Ze staan bij enkele soorten alleen, maar meestal staan ze gegroepeerd, samen met steriele blaadjes of parafysen in een androecium. In de buik van het antheridium worden de zaadcellen (antherozoïden, spermatozoïden) gevormd, die met behulp van twee flagellen kunnen zwemmen.
Het archegonium is min of meer bol, gewoonlijk flesvormig, met een buik en een hals en met een wand van steriele cellen. In de buik bevindt zich een eicel en daarboven een hals met halskanaalcellen. De eicel in de buik van het archegonium wordt bevrucht door een zaadcel uit een antheridium. Deze zaadcel heeft aan een dun vliesje water reeds genoeg om naar het archegonium te zwemmen. De cellen in het halskanaal lossen op en laten de zaadcel door. Ook de archegonia kunnen gegroepeerd staan in een gynoecium, met parafysen tussen de archegonia en beschut door bladen.
- Antheridium, archegonium en jonge sporofyt bij enkele levermossen
-
 Marchantia polymorpha: antheridium en antherozoïden
Marchantia polymorpha: antheridium en antherozoïden -
 Marchantia polymorpha: archegonium
Marchantia polymorpha: archegonium -
 Porella: sporofyt overlangse doorsnede, omhuld door archegoniumrest
Porella: sporofyt overlangse doorsnede, omhuld door archegoniumrest



Bij de bevruchting van de eicel door de zaadcel ontstaat de diploïde zygote. Hieruit ontwikkelt zich eerst een embryo en daaruit groeit de sporofyt op en ten koste van de mosplant (de gametofyt). Op een enkele gametofyt kunnen meerdere sporofyten groeien. De sporofyt bestaat uit een voet, een kapselsteel (seta) en een sporenkapsel (sporangium of sporogoon). Bij levermossen heeft het sporenkapsel een eenvoudige bouw en heeft geen bladgroen. Bij veel mossen kan de bouw zeer complex zijn met chloroplasten in de cellen, huidmondjes en een centrale streng die lijkt op een vaatbundel.
Er zijn enkele groepen van (blad-)mossen (zoals Dicranum en Garovaglia), die heterospoor zijn met grote, vrouwelijke macrosporen en kleine, mannelijke microsporen. Bij sommige soorten leveren de microsporen de dwergmannetjes (kleine mannelijke planten), die op de vrouwelijke plant kunnen groeien.
Varens bewerken
| | |
|
Bij varens[14] is er een afwisseling is van twee morfologisch verschillend gebouwde generaties, waarbij de sporofyt dominant is. De korter levende gametofyt is minder goed ontwikkeld, maar kan bij veel soorten zelfstandig leven. Dit type levenscyclus wordt een heteromorfe, digenetische cyclus met dominante sporofyt genoemd.
Veel varens zijn homospoor en eenhuizig, maar er zijn enkele groepen die heterospoor zijn: de kleinere microsporen worden gevormd in een microsporangium en de grotere macrosporen worden gevormd in een macrosporangium. Microsporangia en macrosporangia worden dan gevonden op sporofyllen of op stegofyllen aan dezelfde, eenhuizige varenplant.
De door meiose gevormde sporen ontwikkelen zich bij isospore varens één type tot de voorkiem of prothallium, maar bij heterospore varens het microprothallium en het macroprothallium. De voorkiem is bij een aantal varensoorten vrijlevend, maar ontwikkelt zich bij sommige andere varensoorten zelfs binnen de beschermende sporenwand. De archegonia en antheridia worden gevormd op, of soms in, de voorkiem.
Er is slechts een onbeweeglijke eicel in de buik van het archegonium. De zaadcellen of spermatozoïden, die flagellen hebben, hebben voor de bevruchting van de eicel water nodig om naar het archegonium te zwemmen.
Na de bevruchting ontwikkelt de zygote zich, via een embryostadium, waar al de aanleg van een echte wortel herkenbaar is, tot een "varenplant", de op zichzelf staande sporofyt, terwijl de prothallium afsterft. De sporofyt is aanzienlijk robuuster en leeft langer dan het prothallium.
- Voortplantingsorganen bij varens en varenachtigen
-
 Gametofyt bij varens:
Gametofyt bij varens:
A-gametofyten, B-archegonia,
C-rizoïden, D-antheridia. -
 Selaginella: microspore met microprothallium (v=vegetatieve cel), antheridium (p, c=antheridiumwand) en spermatozoïden (a).
Selaginella: microspore met microprothallium (v=vegetatieve cel), antheridium (p, c=antheridiumwand) en spermatozoïden (a). -
 Selaginella: macrospore met rizoïden en binnen de sporewand (spm) een macroprothallium (pr) met onbevrucht archegonium (ar) en met 2 embryo's (emb1 en emb2)
Selaginella: macrospore met rizoïden en binnen de sporewand (spm) een macroprothallium (pr) met onbevrucht archegonium (ar) en met 2 embryo's (emb1 en emb2)



Voorbeelden van isospore varens zijn de wolfsklauwen (Lycopodium) en de eikvarens (Polypodium). Voorbeelden van heterospore varens zijn kroosvarens (Azolla) en Selaginella. Bij Selaginella ontwikkelt de macrogametofyt zich zelfs binnen de celwand van de macrospore, en daar vindt ook de bevruchting plaats. Binnen de macrosporewand begint het jonge embryo zich dan te ontwikkelen.
Zaadplanten bewerken
| zaad | → | verspreiding, dormancy, kieming |
→ | kiemplant |
| ↑ | ↓ | |||
| generatieve voortplanting |
groei | |||
| ↑ | ↙ |
ontwikkeling van voortplantingsorganen |
↖ |
↓ |
| voortplantings- stadium |
jeugd- stadium | |||
| ↓ | ↘ |
vegetatieve voortplanting, en verjonging |
↗ |
|
| veroudering | ||||
| ↓ | ||||
| ouderdoms- stadium |
De levenscyclus bij zaadplanten loopt van zaad tot zaad.[15] Hoewel de levenscyclus van zaadplanten gewoonlijk anders wordt beschreven dan in termen van kernfasewisseling en generatiewisseling, volgt hieronder een met de overige Embryophyta vergelijkende beschrijving.
De zaadplanten of Spermatophyta hebben een heteromorfe, digenetische cyclus met dominante sporofyt: van de twee ongelijk gebouwde generaties is de diploïde sporofyt dominant. De sporofyt begint zijn leven als een embryo dat gewoonlijk afhankelijk is van reservevoedsel in het zaad. Het embryo groeit uit via een kiemplantstadium tot een volwassen plant. De gametofyten zijn kortlevend en van microscopische afmetingen.
Bij de zaadplanten zijn er globaal twee grote groepen te onderscheiden: de naaktzadigen (Gymnospermae) en de bedektzadigen (Angiospermae) met daarin de bloemplanten. De terminologie die in verband met de voortplantingsstructuren gebruikt wordt bij de bedektzadigen of bij de bloemplanten kan meestal ook gebruikt worden bij de naaktzadigen, maar soms is het duidelijker ook de termen te gebruiken die ook van toepassing zijn op de varens en op de alleen fossiel bekende zaadvarens of deze naast elkaar te zetten.
Alle zaadplanten zijn heterospoor en heterosporangiaat: de microsporen en macrosporen worden gevormd in van elkaar verschillende sporangia: in het microsporangium, resp. in het macrosporangium. Terwijl bij varens de macrosporen groter zijn dan de microsporen -vandaar de namen-, zijn bij veel zaadplanten de macrosporen veel kleiner dan de microsporen en slechts met een microscoop zichtbaar.
De microsporangia van de zaadplanten groeien aan een structuur die men aanduidt met de term microsporofyl, maar bij de Gnetales en bij de bedektzadigen worden ze traditioneel meeldraad genoemd. De structuren die de microsporangiën dragen (microsporofyllen en meeldraden) zijn gevarieerd van vorm: bladachtig of schubachtig of asjes (al of niet vertakte stengeltjes). Bij de naaktzadigen staan deze microsporofyllen gegroepeerd in een strobilus (sporenaar, kegel). Bij bedektzadigen staan de meeldraden in kransen in bloemen.
De microgametofyt ontwikkelt zich binnen de wand van de microspore en vormt daar een tot een of enkele cellen gereduceerd microprothallium met antheridium; gewoonlijk spreek men nu van pollen.
- Microsporangia en meeldraden
-
 Pinus (den):
Pinus (den):
mannelijke strobilus met groot aantal microsporofyllen en microsporangia. -
 Ephedra (zeedruif):
Ephedra (zeedruif):
mannelijke strobilus met microsporofyllen (meeldraden) en microsporangia. -
 Bedektzadigen meeldraad:
Bedektzadigen meeldraad:
A dwarsdoorsnede met 4 helmhokjes met bij 1 microsporemoedercellen, bij 2 vorming van microsporen en bij 3 rijpe stuifmeelkorrels; k=helmbindsel; s=naad waarlangs het helmhokje openbarst. -
 Pollenkorrels
Pollenkorrels
A rudimentair prothallium enkele kleine cellen y;
B gekiemde pollenkorrel met pollenbuis.




In het macrosporangium worden door macrosporemoedercellen de macrosporen gevormd. Het macrosporangium heet bij de zaadplanten nucellus. De nucellus is omgeven door een of twee beschermende integumenten (ook wel: zaadvliezen). Samen vormen de nucellus en integumenten de zaadknop. De integumenten laten aan de top van de nucellus de micropyle (een kleine opening) vrij, waardoor eventueel de spermatozoïde naar de eicel kan zwemmen.
- Zaaknop met nucellus (macrosporangium)
-
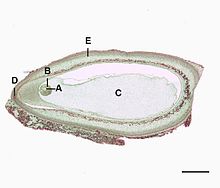 Pinus zaadknop overlangse doorsnede (0.6mm) A=eicel, B=archegonium, C=gametofyt, D=micropyle, E=integument
Pinus zaadknop overlangse doorsnede (0.6mm) A=eicel, B=archegonium, C=gametofyt, D=micropyle, E=integument -
 Ginkgo biloba, E. gesteelde zaadknoppen (rechts)
Ginkgo biloba, E. gesteelde zaadknoppen (rechts) -
 Zaadknop bij zaadplanten
Zaadknop bij zaadplanten -

_(14775959344).jpg)


In de nucellus wordt een of enkele macrosporemoedercellen (ook wel: embryozakmoedercel) gevormd. Een of enkele macrosporemoedercellen ondergaan een reductiedeling of meiose, waarbij elk vier op één lijn liggende, haploïde macrosporen worden gevormd. Van deze vier macrosporen gaan in het algemeen 3 te gronde en blijft de macrospore behouden die het dichtst bij de micropyle ligt: de embryozak.
Bij de naaktzadigen staan de zaadknoppen op vertakte blad- of stengelachtige structuren of op schubben (ook wel: kegelschubben), maar bij de bedektzadigen staan een of meerdere zaadknoppen op een carpel of vruchtblad of worden er door omgeven. De carpellen worden gehomologiseerd met macrosporofyllen.
Naaktzadigen bewerken
De groep naaktzadigen (Gymnospermae) omvat een aantal ordes, waarvan veel alleen fossiel bekend zijn, zoals zaadvarens (Pteridospermales), de Cycadeoidales, de Pentoxylales en de Cordaitales. De groepen met recente vertegenwoordigers zijn de palmvarens, de Ginkgoales, de Coniferales en de Gnetales. Bij deze ordes is er, naast morfologisch onderzoek, ook cytologisch en biochemisch onderzoek mogelijk ten behoeve van het opstellen van een, weliswaar incomplete, fylogenetische stamboom. De slechts fossiel bekende groepen kunnen alleen worden ingedeeld en tussengevoegd op grond van hun bewaard gebleven morfologische kenmerken, in vergelijkend onderzoek met de recente groepen, waarvan de fylogenetische samenhang niet geheel duidelijk is.
Palmvarens bewerken
|
De Cycadales zijn een orde in de klasse Cycadopsida. Cycadales zijn betrekkelijk kleine struiken, bomen of klimplanten met manoxyl secundair hout. De volwassen planten zijn strikt tweehuizig, en hebben een soms knolvormige stam. De macrofyllen of bladen zijn enkel of dubbel veerdelig, waardoor veel planten een palmachtig uiterlijk hebben.[16][17]
De microsporofyllen dragen enkele tientallen tot honderden, in groepen bijeenstaande microsporangia (sori). De tot pollen ontwikkelde microsporen worden door de wind verspreid. De microspore ontwikkelt zich ten tijde van de bestuiving binnen de sporewand tot 3-cellige gametofyt: een prothalliumcel, een generatieve cel en een pollenbuiscel (de grootste cel). Na de bestuiving ontwikkelt de pollenkorrel zich verder in de pollenkamer door de vorming van een pollenbuis die bij de nucellus naar binnen groeit. De generatieve cel deelt zich in een pollenbuiscel (steelcel) en een spermatogene cel. De mannelijke gametofyt bestaat uit een pollenbuis en heeft geen antheridia, maar vormt uit de spermatogene cel twee ongeveer ¼ mm grote spermatozoïden (zaadcellen), die in spiralen staande banden van korte flagellen of cilia hebben.
De spermatozoïde zwemt naar de eicel en bevrucht deze (oogamie). De zygote groeit uit tot een in het zaad gelegen embryo.
De sporangia staan op sporofyllen. De macrosporangia staan op schubachtige macrosporofyllen in eindstandige of axillaire strobili. Bij Cycas vormen de vrouwelijke planten echter geen strobili: de enigszins op macrofyllen gelijkende macrosporofyllen staan aan de top van de stam. Bij sommige Cycas-soorten hebben deze bladachtige macrosporofyllen nog een duidelijke veernervige bladvorm, bij andere soorten zijn ze sterk gereduceerd tot schubben en is de bladvorm niet meer herkenbaar.
De zaadknoppen of ovula (enkelvoud: ovulum) zijn groot en met het blote oog goed zichtbaar. De zaadknoppen staan per 2 (tot 12 bij Cycas) op een macrosporofyl, maar bij sommige groepen zijn één of meer zaadknoppen omgeven door een napje (cupula).
De zaadknop heeft 1 dik integument, dat zich ontwikkelt tot een beschermende, drielagige "testa". Aan het uiteinde heeft de zaadknop een opening, de micropyle.
In de nucellus worden door meiose tetraden van macrosporen zonder tussenwanden gevormd. De macrospore in de zaadknop ontwikkelt zich zo tot macroprothallium (het primair endosperm ), dat eerst vrije kernen heeft (een "coenocytisch weefsel"), maar later celwanden ontwikkelt. Boven het macroprothallium vormt zich een prothalliumkamer, waar ruimte is voor de spermatozoïden. Het macroprothallium heeft twee tot enkele eenvoudig gebouwde archegonia, bestaande uit een archegoniumwand met een eicel en 2 halskanaalcellen.
- voortplantingsorganen bij palmvarens
-
 (Vrouwelijke) macrosporofyl van Dioon edule met 2 zaadknoppen of zaden.
(Vrouwelijke) macrosporofyl van Dioon edule met 2 zaadknoppen of zaden. -
 Macrosporofyllen van Cycas revoluta. Deze staan langs de stam.
Macrosporofyllen van Cycas revoluta. Deze staan langs de stam. -
 Microsporofyllen van Encephalartos villosus. De microsporangia (= pollenzakken) zijn gedeeltelijk geopend.
Microsporofyllen van Encephalartos villosus. De microsporangia (= pollenzakken) zijn gedeeltelijk geopend.



In de zaadknop ontstaat er een pollenkamer doordat de cellen aan de top van de nucellus degenereren. Op de micropyle wordt een kleverige pollinatiedruppel gevormd, waarin het pollen kan blijven hangen. Door indrogen van de pollinatiedruppel die uit de micropyle treedt, wordt het pollen naar binnen gezogen naar de pollenkamer.
De sporofyten van palmvarens (Cycadales) beginnen als een embryo met twee zaadlobben. Het embryo is door een lange suspensor (of "kiemdrager") in de macrogametofyt ( primair endosperm ) geduwd. Een suspensor is een cel die door zijn lengtegroei het embryo in het weefsel van de gametofyt duwt, wat dient als reservevoedsel.
Ginkgoales bewerken
|
De Ginkgoales zijn een orde in de klasse Coniferopsida.[20] Er is slecht 1 recente soort, Ginkgo biloba (Japanse notenboom) in de familie Ginkgoaceae, die mogelijk wild voorkomt in China. Het zijn strikt tweehuizige bomen, met lange en korte loten. De bladen hebben dichotoom vertakte bladnerven. De bladeren vertonen bladdimorfie: aan de lange loten zijn de bladen wat dieper ingesneden dan die aan de korte loten. Het embryo heeft twee kiemlobben en geen duidelijke suspensor.
De microsporangia staan in strobili met zijasjes (ook wel sporangioforen) die elk aan de vebrede top 2 microsporangia dragen. De ontwikkeling van de microspore bij Ginkgo biloba vertoont een sterke overeenkomst met die bij de den (Pinus).[21] De microgametofyt vormt twee 2 prothalliumcellen, een pollenbuiskern en 2 spermatozoïden die een spiraalband met trilharen hebben.
Aan de vrouwelijke plant vindt men de eindstandige ovula (zaadknoppen) in paren op een dichotoom vertakte as. Aan de voet van de zaadknop bevindt zich een verbrede ring, de cupula. Binnen de integumenten heeft de nucellus aan de top een pollenkamer. Door meiose ontstaat er een lineaire sporentetrade van macrosporen. De onderste daarvan ontwikkelt zich tot een macroprothallium (gametofyt). Deze ontstaat door vrije delingen van de celkern tot er ongeveer 256 kernen zijn. Hierna worden pas de tussenliggende celwanden gevormd. In het chlorofyl bevattende prothallium groeien twee tot drie archegonia, die elk een eicel , een ventrale kanaalcel en twee tot vier halscellen bevatten. Boven het archegonium ontstaat een ringvormige archegoniumkamer. Het macroprothallium vormt zetmeelrijk haploïde primair endosperm .
- Japanse notenboom
-
 mannelijke kegels
mannelijke kegels -
 vrouwelijke bloeiwijzen
vrouwelijke bloeiwijzen -
 zaden
zaden



De voortplantingsstructuren van Ginkgoales en de Cycadales komen (door convergente evolutie) in veel kenmerken met elkaar overeen, zoals het bestuivingsmechanisme, de bouw van de microgametofyt en van de microgameten, de zaadknoppen en het zaad. Het zaad heeft een harde binnenlaag en een vlezige buitenlaag. Het embryo heeft 2 kiembladen.
Coniferen bewerken
|
De Coniferales zijn gewoonlijk forse bomen met pycnoxyl secundair hout, dat wil zeggen: massief hout met smalle mergstralen. De naaldvormige bladen zijn primair macrofyllen met dichtoom vertakte nerven, maar secundair 1-nervige microfyllen.[23]
De voortplantingsorganen van de Coniferales (coniferen) zijn in de verschillende groepen zeer verschillend in bouw. De kegels met de sporangia zijn eenslachtig.
De vrouwelijke kegels zijn samengesteld: er is een hoofdas met vele tot enkele schubvormige schutbladen. De zaadknoppen-dragende schub kan daarmee in mindere of meerdere mate vergroeid zijn. De zaadknoppen bestaat uit een nucellus met 1 integument. De daarin aanwezige macrosporemoedercellen vormen door meiose één functionele macrospore, de overige drie macrosporen degenereren. De macrospore groeit uit via een coenocytisch stadium het macroprothallium met (1-) 3 (-5) archegonia.
Bij Pinus (den) heeft de mannelijke strobilus schubvormig microsporofyllen met 2-15 microsporangia of pollenzakjes aan een schub.
De microsporemoedercellen (microsporocyten) vormen door meiose de microsporen, die blaasvormige uitstulpingen hebben. Het microprothallium bij de families Pinaceae, Podocarpaceae, Araucariaceae van de Coniferales bestaat uit microprothalliumcellen en een pollenbuiscel, waarnaast de generatieve cel nog 2 kernen vormt: een steriele cel (soms "steelcel" genoemd) en een spermatogene cel. Uit deze laatste worden de mannelijke gameten gevormd.
Bij Cephalotaxaceae, Cupressaceae, inclusief Taxodiaceae gaat de ontwikkeling, na de vorming van de kern van de microspore, direct verder met de vorming van de pollenbuiskern en de generative cel.[24]
- Coniferales
-
 Juniperus communis (jeneverbes),
Juniperus communis (jeneverbes),
tak met vruchten -
 Taxus baccata (venijnboom, taxus),
Taxus baccata (venijnboom, taxus),
detail takje met vruchten -
 Podocarpus polystachys,
Podocarpus polystachys,
bladeren -
 Cephalotaxus harringtonia,
Cephalotaxus harringtonia,
takken met vruchten -
 Pinus sylvestris (grove den),
Pinus sylvestris (grove den),
lang lot met 2 zaadkegels tussen bebladerde korte loten -
 Larix decidua (Europese lork),
Larix decidua (Europese lork),
takje met bebladerde korte loten -
 Taxodium distichum (moerascipres),
Taxodium distichum (moerascipres),
in natuurlijke omgeving
.JPG)






De bevruchting vindt plaats door windbestuiving en sifonogamie: de pollenbuis groeit naar de eicel , waar de spermakern versmelt met de eicel . Er vinden meer bevruchtingen plaats, zodat er meer embryo's ontstaan. Het vormen van meerdere embryo's heet polyembryonie. De kiemplanten hebben tot 15 zaadlobben (cotylen).
Gnetales bewerken

groen – Welwitschia
blauw – Gnetum
rood – Ephedra
purper – Gnetum en Ephedra overlap verspreidingsgebied
De Gnetales is een tot de zaadplanten behorende orde en aan de naaktzadigen verwante planten. Tot de orde Gnetales behoren drie geslachten, elk in een eigen familie: de Gnetaceae met Gnetum, de Ephedraceae met Ephedra en de Welwitschiaceae met Welwitschia.[25] De oude naam Chlamydospermae (mantelzadigen) werd ook gebruikt voor dit taxon, dat dan naast de naaktzadigen en de bedektzadigen werd geplaatst.
De Gnetales zijn houtige planten met secundair hout zonder harskanalen. De bladeren zijn tegenoverstaand. Belangrijke kenmerken van de orde zijn de houtvaten (tracheeën) in het secundaire hout, waarschijnlijk door parallelle evolutie uit tracheïden. De planten zijn gewoonlijk tweehuizig met meestal eenslachtig eenslachtige "bloemen". De voortplantingsorganen ("bloemen") staan gegroepeerd in samengestelde "bloeiwijzen" (strobili of kegels). Het embryo heeft twee kiembladen (cotylen).
De mannelijke "bloem" bestaat uit een "periant" (bloemdek) met daarbinnen antheroforen met elk 2 tot 8 synangia (ook wel "pollenzakjes") - een tot een samenhangend geheel vergroeide sporangia.
De vrouwelijke "bloemen" hebben een enkele, rechtopstaande ovulum, waarvan de nucellus is omgeven door twee of drie omhulsels: twee integumenten en een "periant".
Een micropyle is de opening van het integument dat de nucellus (macrosporangium) omgeeft. Een opvallend kenmerk is de lang uittredende micropylaire buis.
In de nucellus vormen zich door meiotische deling uit een sporemoedercel de vier haploïde macrosporen in een tetrade. Hiervan gaan er drie te gronde. De overgebleven macrospore (of tenminste de kern van de macrospore) deelt zich meermaals tot enkele of meerdere cellen of celkernen: het macroprothallium of macrogametofyt. Ook dit wordt wel "embryozak" genoemd. Een van de celkernen van de embryozak functioneert als eicel.
De micropyle groeit bij de Gnetales uit tot een lang uittredende micropylaire buis, waardoorheen een druppel vocht naar buiten wordt gevormd om de stuifmeelkorrels in de lucht op te kunnen vangen. Bij indrogen van het vocht worden de stuifmeelkorrels naar binnen gezogen. Deze wijze van bestuiving wordt het "druppelmechanisme" genoemd. De bevruchting vindt plaats door de vorming van een pollenbuis met twee mannelijke gameetkernen.
Gnetaceae bewerken
|
De Gnetaceae zijn een familie van naaktzadigen. Meestal wordt aangenomen dat het bestaat uit één geslacht, Gnetum, maar soms worden twee geslachten geaccepteerd.
Gnetum heeft tegenover elkaar geplaatste, grote netnervige bladen. De bomen hebben een normale secundaire diktegroei. Anders dan bij de gymnospermen, bevat bij sommige soorten het secundaire floëem, naast de zeefvaten, ook begeleidende cellen. Naast houtvaten zijn er ook tracheïden. Sommige soorten zijn tweehuizig, maar bij andere is er soms tussen de meeldraden een, meestal steriele, zaadknop te vinden. Van Gnetum gnemon (melindjoe) is de voortplanting het best onderzocht en wordt dan ook hier besproken.
De "bloemen" staan in kransen op kegels (strobuli). De kegels staan meestal in de oksel van een blad of aan het uiteinde van een dwergloot. Een strobilus bestaat uit een as met boven elkaar geplaatste kragen, waarboven de kransen mannelijke of vrouwelijke bloemen staan. Twee aan de voet ervan geplaatste bracteeën (schutbladen) omgeven de jonge strobilus.
Op de mannelijke planten vindt men de mannelijke strobili of "bloeiwijzen" met, binnen een periant, de meeldraden, de microsporofyllen met microsporangia.
De mannelijke strobilus, die vertakt kan zijn, heeft boven de kraag verschillende kransen met mannelijke "bloemen".
Een mannelijke "bloem" wordt gevormd door een antherofoor of microsporangiofoor, die een synangium (enkele met elkaar vergroeide sporangiën) draagt. Dit geheel wordt ook wel "meeldraad" genoemd. Aan de basis hiervan staan twee tot een periant vergroeide bracteolen. Naast dergelijke bloemen met slechts een meeldraad zijn er ook met meer dan één meeldraad. Soms is er een gereduceerde, steriele zaadknop aanwezig.
De microsporen worden gevormd door meiotische delingen de microsporemoedercellen in de microsporangia (pollenzakjes) van het synangium op de sporangiofoor. Ten tijde van de bestuiving bevat de pollenkorrel drie kernen: een pollenbuiskern, een steriele cel en een spermatogene cel. Uit de spermatogene cel vormen zich twee niet-beweeglijke mannelijke cellen.
Vrouwelijke bloemen staan in kransen in een aarachtige bloeiwijze, en worden ondersteund door een kraag. De vrouwelijke bloem bestaat uit een nucellus, die door drie omhulsels wordt omgeven: van buiten naar binnen een periant en twee integumenten, waarvan de binnenste integument uitsteekt als een micropylaire buis.
In de nucellus worden uit archesporecellen 8-10 macrosporemoedercellen gevormd. Door meiose wordt een celkern van een macrosporetetrade zonder celwanden gevormd. Hieruit groeit door mitotische celdelingen, ook weer zonder vorming van celwanden, een embryozak (prothallium). Er worden geen archegonia gevormd en een van de kernen van het prothallium fungeert dan als eicelkern.
-
 botanische afbeelding
botanische afbeelding -
 Gnetum latifolium var. funiculare
Gnetum latifolium var. funiculare
mannelijke bloemen -
 Gnetum gnemon
Gnetum gnemon
vrouwelijke bloemen (enkele met micropylaire buis) en vruchten
_(20947583061).jpg)

_(6863727669).jpg)
Het door de wind verspreide stuifmeel wordt opgevangen door een pollinatiedruppel op de uitstekende micropylaire buis. De bevruchting vindt plaats door sifonogamie : meerdere pollenbuizen dringen door in het prothallium, waarvan meerdere kernen bevrucht worden. Hierna worden pas de celwanden gevormd.
Elke van de zygotes deelt zich en vormt twee primaire suspensorbuizen. Bij verdere groei vormen zich aan de top de meercellige secundaire suspensor. Aan het uiteinde daarvan ontwikkelt zich echte embryo's. Gnetum vertoont op deze manier een extreme vorm van polyembryonie. In het uiteindelijke zaad blijft er slechts een embryo over. Het embryo heeft zijdelings in het zaad uitstekende uitgroeisels, waarmee het embryo zich kan voeden. Dit verschijnsel komt verder alleen nog voor bij Welwitschia.
Welwitschiaceae bewerken
|
Welwitschia mirabilis,[26][27][28] synoniem: W. bainesii, is de enige soort die wordt gerekend tot de familie Welwitschiaceae, een monotypische familie met slechts één geslacht en slechts één soort. De plant is een woestijnplant en komt voor in de kuststreken van Angola en Namibië. De plant is tweehuizig: er zijn aparte mannelijke en vrouwelijke planten. De "bloemen" staan in een strobili, die in de oksel van de twee tegenoverstaande bladeren staan.
De mannelijke bloemen zijn omgeven door twee paar kruiswijs tegenoverstaande, vergroeide bracteeën. De mannelijke bloemen bevat in het midden een door rudimentaire schubben omgeven gereduceerde zaadknop. Deze zaadknop produceert ongeveer voor 50% uit suikers bestaande nectar. De zes onderaan hun basis vergroeide meeldraden (ook wel sporangiofoor genoemd) staan in een krans. Elk meeldraad draagt een synangium van drie met elkaar vergroeide microsporangia of pollenzakken. De stuifmeelzak opent in de vorm van vaak korte sleuven, net als bij de andere vertegenwoordigers van Gnetopsida. De stuifmeelkorrels worden door de uitdrogende pollenzak naar buiten geperst.
De mannelijke gametofyt bestaat uit spermatogene cel naast twee andere cellen. De spermatogene cel deelt zich in twee spermacellen.
De vrouwelijke bloemen zijn omgeven door twee paren vergroeide bracteeën. In de zaden rijpen het binnenste paar bracteeën is moeilijk, de uiterlijke vormen vleugels. Elke bloem bevat een rechtopstaande zaadknop. Het integument is een micropylaire buis, waardoor een pollinatiedruppel of nectar wordt uitgescheiden.
De vrouwelijke gametofyt ontstaat door vrije nucleaire delingen van alle vier door meiose ontstane kernen en daaropvolgende vorming celwand. De gametofyt kan uit tot duizend cellen bestaan. Er worden geen archegonia gevormd, en ook de eicel is niet te onderscheiden van de andere archegoniumcellen. Met een prothallium-buis groeit de gametofyt de pollenbuis tegemoet.
- Welwitschia mirabilis
-
 Verspreidingsgebied van Welwitschia mirabilis.
Verspreidingsgebied van Welwitschia mirabilis. -
 bloemdiagram van een vrouwelijke bloem
bloemdiagram van een vrouwelijke bloem -
 bloemdiagram van een mannelijke bloem
bloemdiagram van een mannelijke bloem -
 Welwitschia mirabilis
Welwitschia mirabilis -
 De bloeikegels van een Welwitschia met rode Welwitschiawantsen.
De bloeikegels van een Welwitschia met rode Welwitschiawantsen. -
 Vrouwelijke kegels / bloeiwijzen
Vrouwelijke kegels / bloeiwijzen -
 mannelijke kegels / bloeiwijzen
mannelijke kegels / bloeiwijzen -
 Welwitschia mirabilis
Welwitschia mirabilis








De plant bloeit van midzomer tot de herfst. De bloemen worden bestoven door insecten, mogelijk (mede) door de welwitschiawants. De wants Probergrothius sexpunctatis voedt zich met de nectar, maar bestuiving is nog niet duidelijk aangetoond. De bevruchting vindt plaats via een pollenbuis (siphonogamie).
De zaden hebben een embryo met een worteltje en twee zaadlobben. Ze rijpen in de lente en vrijgegeven door verval van de as van de kegel. De zaden zijn ongeveer 3,5 × 2,5 cm groot en hebben een papierachtige vleugel. De wind zorgt voor de verspreiding van de zaden. De zaden blijven enkele jaren kiemkrachtig. Van de ongeveer 10.000 tot 20.000 bloemen per plant en jaar plaats slechts ongeveer 20 tot 200 levensvatbare zaden. Ze ontkiemen pas na zware regenval.
Ephedraceae bewerken
|
De meeste soorten van het geslacht Ephedra zijn tweehuizig. Het zijn sterk vertakte struiken met groene takken die lang dun zijn. De kruiswijs tegenoverstaande bladeren zijn schubvormig. De "bloemen" staan in "aren" of strobuli aan het eind van korte zijtakken.
De mannelijke strobilus heeft structuren die "bloemen" genoemd kunnen worden. Ze staan in de oksel van kruiswijs tegenoverstaande schutbladen. De "meeldraad" (ook wel: antheroforen of microsporangioforen) staan binnen een periant en bestaat uit een asje met aan het uiteinde enkele microsporangia (pollenzak).
Het microsporangium bevat de microsporemoedercellen, die de microsporen vormen. Bij de meiose worden tetraden van microsporen gevormd, die zich verder ontwikkelen door gewone celdelingen. Daarbij ontstaan 2 prothalliumcellen, een generatieve cel en een pollenbuiskern. De generatieve cel deelt zich in een steriele cel ("steelcel") en een spermatogene cel.
De vrouwelijke "bloemen" bestaan uit een strobilus met een eindstandige zaadknop, die op een asje staat met enkele paren kruiswijs tegenoverstaand schutbladen.
De zaadknop bestaat uit een nucellus (macrosporangium) met twee integumenten. Van de zaadknop is de micropyle van het binnenste integument uitgegroeid tot een micropylaire buis. De integumenten laten bovenaan bij de nucellus een ruimte vrij: de pollenkamer.
In de nucellus ligt een macrosporemoedercel, die door meiose een tetrade van macrosporen vormt - daarvan gaan er drie te gronde en slechts één macrogametofyt met enkele archegonia ontwikkelt zich uit de overgebleven macrospore. In het archegonium liggen de eikern en daarboven de ventrale kanaalcel.
Door middel van het druppelmechanisme worden door de wind verspreide pollenkorrels ingevangen.
-
 Ephedra distachya, botanische tekening
Ephedra distachya, botanische tekening -
 Ephedra foeminea, bloemen met meeldraden en zaadknoppen (met pollinatiedruppel)
Ephedra foeminea, bloemen met meeldraden en zaadknoppen (met pollinatiedruppel) -
 Ephedra distachya, vrouwelijke bloemen
Ephedra distachya, vrouwelijke bloemen -
 Ephedra fragilis, rijpe zaden, met gekleurde schutbladen
Ephedra fragilis, rijpe zaden, met gekleurde schutbladen


.jpg)

In het stadium, waarin de pollenkorrel 2 prothalliumcellen, een spermatogene cel, een steriele cel en een pollenbuiskern heeft, vindt de pollinisatie of bestuiving plaats. Door het indrogen van de pollinatiedruppen komt de stuifmeelkorrel in de pollenkamer, waar de pollenbuis uitgroeit door het nucellusweefsel in het archegonium. De spermatogene cel deelt zich in twee spermakernen. Er vindt bevruchting plaats van de eickern en dubbele bevruchting van de ventrale kanaalcel van het archegonium.
Bedektzadigen, Angiospermae bewerken
|
Bij bedektzadigen (Angiospermae) zijn de voortplantingsorganen gewoonlijk verzameld in bloemen. Een bloem is een ingewikkelde voortplantingsstructuur, waarin grote verscheidenheid bestaat in de bouw. Een volledige, eenhuizige bloem bestaat uit een bloembodem, een kelk, een bloemkroon, meeldraden en een stamper met vruchtbeginsel, stijl en stempel. De vrouwelijke voortplantingsorganen zijn de in een gesloten vruchtbeginsel gegroepeerde zaadknoppen of zaadbeginsels of ovula). De mannelijke voortplantingsorganen zijn de meeldraden met helmdraad (filament) en een helmknop (anthere). De ontwikkeling van de gametofyten vindt plaats binnen de zaadknoppen en de stuimeelkorrels.[30]
Stamper, vruchtbeginsel en zaadknoppen bewerken

· Grijs: Bloemsteel en bloemboden
· Donkergrijs: Kelkbladen
· Lichtgrijs: Kroonbladen
· Zwart: Krans van meeldraden
· Rood: Stamper
De stamper (pistillum, ovarium) is een onderdeel van een vrouwelijke of van een tweeslachtige bloem, dat aan het uiteinde van de bloemas staat.
Een volledige bloem bestaat (van beneden naar boven) uit de bloemsteel, de bloembodem; daarop ingeplant de kelk (met de kelkbladen), de kroon (met de kroonbladen), een krans van meeldraden en de stamper. Soms is de stamper gedeeltelijk of geheel verzonken in de bloembodem (die dus de stamper dan omgeeft).
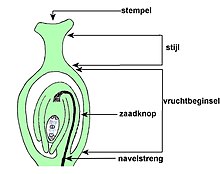
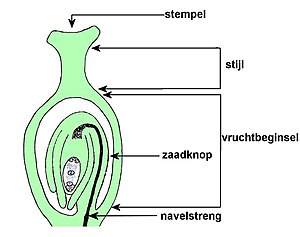
De stamper is opgebouwd uit vruchtbeginsel, stijl en stempel. Een enkelvoudige stamper is opgebouwd uit één vruchtblad, maar een samengestelde stamper is uit meer vruchtbladen opgebouwd.[31] Het vruchtbeginsel is het onderste gedeelte van de stamper en bevat de zaadknoppen (ovulum).
Vruchtbeginsel en stijl bewerken
Het vruchtbeginsel bestaat uit één of meer samengevouwen vruchtbladen, die bij de rand en/of met elkaar vergroeid zijn. Bij een apocarp vruchtbeginsel zijn de vruchtbladen niet met elkaar vergroeid. Daartegenover staat het coenocarpe vruchtbeginsel, waar de vruchtbladen aan elkaar vergroeid zijn. De vruchtbladen zijn vaak niet meer als blad herkenbaar. Het gesloten vruchtbeginsel van de bedektzadigen bevat de niet-vrijliggende zaadknoppen.
De vruchtbladen vormen één of meer holten (eenhokkig, tweehokkig, driehokkig tot veelhokkig). De holten zijn daarbij gescheiden door tussenschotten, die tot het midden doorlopen. Het vruchtbeginsel blijft eenhokkig als de tussenschotten niet geheel tot het midden doorlopen.
De stijl is het gedeelte dat op het vruchtbeginsel staat of tussen de delen van het vruchtbeginsel. De stijl is meestal langgerekt. Er kan meer dan één stijl zijn (bijvoorbeeld voor elk vruchtblad één), die vrij staan of min of meer vergroeid. Binnen een soort kan er een variatie zijn in de lengte van de stijlen: bij "heterodistylie" zijn er bloemen met lange en andere met korte stijlen; bij "heterotristylie" zijn er bloemen met stijlen in drie verschillende lengten. De stempel staat boven aan de stijl. Ten tijde van de rijpheid is het kleverig en dient voor het opvangen van het stuifmeel. Het stuifmeel wordt door de wind of door insecten gebracht. De stijlen leiden naar het vruchtbeginsel. Na de bestuiving groeien de pollenkorrels uit door de stijl naar de zaadknoppen.
Zaadknop bewerken
De zaadknoppen zijn met een zaadstreng (funiculus of navelstreng) bevestigd aan de zaadlijsten (placentae) van het vruchtbeginsel. De plaats van de zaadlijsten (de aanhechting van de zaadknoppen) in het vruchtbeginsel kan zijn: in de hoeken (hoekstandige placentatie) of aan de buitenwand (pariëtale placentatie), in het midden (centrale placentatie), op de bodem (basale placentatie) of in de top (apicale placentatie) van het vruchtbeginsel.
-
 Enkele typen placentatie: vruchtbeginsels met placenta (lichtgroen) met zaadknoppen (zwart)
Enkele typen placentatie: vruchtbeginsels met placenta (lichtgroen) met zaadknoppen (zwart) -
 Stamper met eenhokkig vruchtbeginsel
Stamper met eenhokkig vruchtbeginsel
met slechts één anatrope zaadknop -
 anatrope zaadknop schematisch
anatrope zaadknop schematisch

De zaadknoppen bevatten een nucellus (megasporangium of zaadknopkern), die door één of twee integumenten (zaadvliezen) wordt omgeven en slechts één functionele macrospore (megaspore) bevat. De zaadknop heeft of één integument en een kleine nucellus, of twee integumenten en een grote nucellus.[32]
Embryozak met eicel, synergiden, antipoden en embryozakkernen bewerken

De macrosporemoedercel (macrosporocyt) ondergaat meiotische delingen, waarbij vier macrosporen worden gevormd. De meiotische deling verloopt asymmetrisch en leidt tot ophoping van cytoplasma in de macrosporemoedercel die de embryozak voortbrengt, de eigenlijke macrogametofyt. De andere dochtercellen gaan te gronde. Een volgroeide embryozak ontstaat door gewone mitotische delingen, verplaatsing van kernen en cytokinese. Hierbij is er een aanzienlijke variatie in de patronen van embryozak-ontwikkeling.
Een veelvoorkomend patroon is, dat er drie mitotische delingen plaatsvinden, waarbij veel soorten dan drie antipoden, twee poolkernen, een eicel en twee synergiden (= steuncellen) worden gevormd. De eicel met beide synergiden bevindt zich aan de micropylaire zijde van de embryozak, de antipoden bevinden zich daartegenover en de poolkernen bevinden zich in het midden.
Meeldraad, helmknop, pollenzak, stuifmeel bewerken


Bij bedektzadigen staan de meeldraden georganiseerd in bloemen. De meeldraden bestaan uit helmdraad (filament) met aan hun top daarop ingeplant de helmknoppen (antheren).[33]
In de helmknoppen met elk twee helmhokken (thecae) bevinden zich in een jong stadium vier strengen met microsporemoedercellen (pollenmoedercellen), twee per helmhok. De strengen vormen zich in de vier pollenzakken (loculi). De vier pollenzakken versmelten later in paren en vormen bij rijpheid twee helmhokken (thecae), die dan gevuld zijn met het pollen.
De binnenwand van de pollenzakken bestaat uit het tapetum, waarbinnen de stuifmeelkorrelmoedercellen (pollenmoederzakken, microsporemoederzakken, microsporocyten) liggen. Het tapetum is een cellaag die zorgt voor de voeding van de er binnen liggende stuifmeelkorrelmoedercellen.
Hier vindt de vorming van stuifmeel- of pollenkorrels plaats. Door meiose ontstaan uit de diploïde microsporemoederzak (microsporocyt) eerst twee cellen (een dyade) en vervolgens een klompje van vier haploïde cellen, een tetrade, van microsporen. De cellen van de tetrade maken zich gewoonlijk van elkaar los en vormen de haploïde microsporen. Dit is in enkele gevallen later nog goed aan de latere pollenkorrels te zien.
Een microsporemoedercel vormt door twee meiotische delingen een groepje van 4 –haploïde– microsporen, (eerst) in tetraden of op een rij, later meestal los. De buitenwand om deze microspore verdikt zich onder invloed van zowel het tapetum als de microspore tot de exine. Daarbinnen wordt door de microspore de intine (binnenwand) gevormd.
In een volgende stap ontwikkelen de microsporen zich door twee gewone mitotische delingen van de celkern binnen de sporenwand. De zich ontwikkelende sporen heten dan stuifmeel- of pollenkorrels. In elke pollenkorrel vormt zich ten tijde van de bestuiving gewoonlijk een grotere vegetatieve en een kleinere generatieve celkern, zodat de pollenkorrel slechts tweecellig is. De vegetatieve cel omsluit de generatieve kern. De generatieve kern heeft een hogere concentratie celplasma om zich heen en vormt na een tweede mitose twee geslachtscellen (gameten), de microgameten of spermacellen, die ieder een eigen kern bevatten. De vorming van de gameten vindt gewoonlijk plaats in de pollenbuis, die van de stempel naar da zaadknop groeit, maar in sommige gevallen reeds voor de bestuiving in de rijpende pollenkorrel.
De microsporen groeien uit tot stuifmeelkorrels doordat de microsporen zich mitotische delen. Uit deze deling ontstaan een vegetatieve en een generatieve cel, ieder met een eigen kern. De vegetatieve cel groeit bij de bestuiving uit tot de pollenbuis. De generatieve cel ondergaat nog een deling waarbij twee spermacellen ontstaan.
Het bovengenoemd verloop komt het meeste voor maar er zijn veel variaties, zodat het niet geldt voor alle bedektzadigen.
De pollenzakjes komen overeen met de microsporangia bij de naaktzadigen en bij de varens. De helmknoppen kunnen gezien worden als een synangium: een verzameling van vergroeide microsporangia. Daarbij zou elke pollenzak een sporangium zijn.
De pollenkorrel bij bedektzadigen (angiospermen) is sterk gereduceerd en homoloog met een microprothallium met antheridium bij de varens en komt dus overeen met een microsporofyt. De reductie gaat bij veel soorten zover, dat er geen tussenwanden meer worden gevormd, en dat het microprothallium en de antheridia niet meer herkenbaar zijn of niet meer worden gevormd.
Bestuiving, bevruchting, zaadvorming bewerken
De bestuiving vindt plaatst als de pollenkorrel op de stempel van de stamper komt. Wanneer de stuifmeelkorrel in aanraking komt met een stempel zwelt deze op doordat osmotisch water wordt opgenomen. Het cytoplasma wordt geactiveerd.
De korrel kiemt door de celwand waarbij er een pollenbuis ontstaat. De pollenbuis maakt een opening in de cuticula van de stempel en begint hierna tussen de cellen van het geleidingsweefsel van de stijl door te groeien in de richting van het zaadbeginsel, waarbij het zich een weg door de stijl baant. Bij sommige planten (lelies) is er geen geleidingsweefsel maar een open stijlkanaal. Tijdens de groei van de pollenbuizen kan er certatie plaatsvinden. Certatie is het verschijnsel dat de stuifmeelbuizen niet allemaal even snel groeien. Zo groeien stuifmeelbuizen van stuifmeelkorrels van nauw verwante planten soms langzamer om inteelt te voorkomen.
De generatieve cel deelt zich mitotische nog een keer in twee generatieve cellen (spermacellen).- De pollenbuis groeit door de kiemopening (micropyle) van het vruchtbeginsel de embryozak in. Dit wordt porogamie genoemd.
Nadat het stuifmeel op de stempel van de stamper is aangekomen, groeit uit de stuifmeelkorrels in de richting van een eicel in de zaadknop een pollenbuis, waardoorheen de gameten (spermakernen) naar de eicel gaan. Dit type bevruchting heet sifonogamie.
In de embryozak (het macroprothallium) versmelten de twee polaire kernen tot een diploïde embryozakkern. De eicel bevindt zich aan de micropyle-zijde van de embryozak, met daarnaast de synergiden, die behulpzaam zijn bij de bevruchting.
In de pollenbuis verplaatsen de twee spermakernen zich naar de embryozak. Er vindt een dubbele bevruchting plaats, dat wil zeggen: een van deze spermakernen versmelt met de kern van eicel. De andere spermakern versmelt met de diploïde secundaire embryozakkern en vormt zo een triploïde (3n) kern. Deze kern groeit uit tot het secundair endosperm (kiemwit) , dat dient als reservevoedsel voor het zich later ontwikkelende embryo.
Op het hierboven beschreven patroon bestaan zeer veel belangrijke variaties.
Zaad en vrucht bewerken
Bij zaadplanten zorgen de zaden en de vruchten voor de verbreiding van de soort. Zaden zijn de bevruchte zaadbeginsels. De vruchten zijn, volgens een morfologische definitie, de uitgegroeide vruchtbeginsels. Deze definitie zou kunnen inhouden, dat bloemen met een onderstandig vruchtbeginsel, schijnvruchten vormen, maar deze strikte opvatting wordt zelden gehanteerd. Volgens een andere, meer ecologische opvatting is, bestaat de vrucht uit de bloem met rijpe zaden. Volgens een nog ruimere opvatting zijn vruchten die structuren die zaden bevatten, zelfs delen van de bloemsteel of zelfs hele bloeiwijzen.[34]
De oorsprong en evolutie van zaden[35] ligt ongeveer 370 miljoen jaar geleden, en begon in de late Devoon. De vroegste zaadplanten komen voort uit een diverse groep van sporevormende planten, de Progymnospermophyta, die uiteindelijk leidden tot zowel de naaktzadigen en angiospermen.
De Progymnospermophyta hadden een varenachtige voortplantingscyclus met vegetatieve structuren die kenmerkend zijn voor zaadplanten. Hun verspreidingseenheden waren sporen die zich ontwikkelden tot gametofyten, waarin de mannelijke en vrouwelijke gameten werden gevormd. Net als varens, werd de bevruchting bij Progymnospermophyta waarschijnlijk mogelijk gemaakt door spermatozoïden, die water nodig hebben voor hun voortbeweging. Daarentegen evolueerden bij zaadplanten pollen als het transportmiddel voor het afleveren van spermacellen bij de vrouwelijke gametofyt, waardoor bevruchting plaatsvindt in afwezigheid van water mogelijk werd.
Zaden bewerken
| Onderdelen van de bloem |
|---|
|
Zaden zijn de na de bevruchting uitgegroeide zaadknoppen (zaadbeginsels). Een zaad bestaat uit een embryo (kiempje), reservevoedsel en een zaadmantel (testa). De zaden ontwikkelen zich bij naaktzadigen de zaadknoppen die vrij staan aan het eind van een asje (takje) of op in kegels staande schubben. Hier is geen sprake van vruchten. Bij bedektzadigen staan de zaadknoppen op vruchtbladen (carpellen) in het vruchtbeginsel, dat de zaadknoppen beschermt.
Zaden bevatten een kiempje of embryo en eventueel ook reservevoedsel voor het kiempje. Het kiempje bestaat uit een pluimpje met een stengeltje en opgevouwen zaadlobben en een worteltje en eventueel enkele bladeren.
Aan het zaad zijn soms nog enkele kenmerken van de zaadknop herkenbaar.[36]
Het zaad wordt omgeven door de testa of zaadhuid, een meer of minder dikke zaadhuid die afkomstig is van de beide integumenten uit het vruchtbeginsel die een beschermende buitenste laag van dode cellen vormen. De zaadhuid bij appel is leerachtig, maar bij braam of rode bes versteend. De zaadhuid kan vergroeid zijn met een deel van de vruchtwand, het pericarp, in welk geval het dan eigenlijk om een vrucht gaat.
De navel (hilum) zaad is een litteken op de plaats waar het zaad met de navelstreng aan de zaadlijst (placenta) was vastgehecht. Soms loopt de navel verder over het zaad als zaadnerf (rafe).
De chalaza of het vaatmerk is de plaats tegenover de navel waar de vaatbundel eindigt.
Verder heeft het zaad een micropyle of poortje (de opening in de integumenten naar de nucellus), waardoor de pollenbuis van de stuifmeelkorrel naar binnen groeit of het worteltje van het kiempje bij de kieming naar buiten komt.
Aan sommige zaden zit een zaadmantel (ook wel: arillus of zaadrok), een vlezig (fruitachtig) omhulsel of aanhangsel. Het gaat hier om een uitgroeisel van het zaad of de zaadsteel.
Het reservevoedsel dat het zaad meekrijgt is opgeslagen in de cellen van het perisperm, het primair of het secundair endosperm of in de zaadlobben van de kiem.
- Opslag van reservevoedsel in het perisperm, dit is afgeleid van het weefsel van de moederplant. Dit komt zelden voor. Het perisperm is diploïde, omdat het gevormd werd door de nucellus (macrosporangium). In basale bedektzadigen (Nymphaeaceae) is het perisperm het belangrijkste voedingsweefsel.
- Opslag van reservevoedsel in het primair endosperm komt bijvoorbeeld voor bij naaktzadigen, maar kwam ook voor bij de uitgestorven zaadvarens. Het primaire endosperm werd gevormd na de meiose en is haploïde: de macrogametofyt. Deze omgeeft het kiempje en is de belangrijkste bron van voedingsstoffen.
- Opslag van reservevoedsel in het secundair endosperm komt veel voor. Het secundaire endosperm werd gevormd bij de dubbele bevruchting uit een polyploïde (meestal triploïde) cel. Het triploïde secundaire endosperm dient als het primaire voedingsweefsel voor het zaad, vaak aangevuld met opslag in de zaadlobben.
- Opslag van reservevoedsel in de zaad- of kiemlobben van de jonge spruit komt bijvoorbeeld voor bij peulvruchten. De zaadlobben van het kiempje en de jonge plant zijn diploïde omdat het kiempje is ontstaan na de bevruchting. Bij de tweezaadlobbigen (dicotylen) is meestal het reservevoedsel uit het endosperm overgegaan in de beide zaadlobben (cotylen) van de kiemplant.
Vruchten bewerken
Vruchten zijn de na de bevruchting de bevruchte zaadknoppen met uitgegroeide stampers bij bedektzadigen, met name de vruchtbeginsels, stijl en stempel. Deze laatste twee kunnen bij veel soorten afvallen. Ook andere delen kunnen echter meegroeien, die men dan ook tot de vrucht rekent.[37] De eerste noemt men dan "echte" vruchten, maar bij de "schijnvruchten" vormen andere delen dan het vruchtbeginsel het hoofdbestanddeel.
Als de vruchtbladen hierbij betrokken zijn, ontstaat een vrucht. Soms zijn nog andere bloemdelen, zoals de bloembodem, betrokken bij deze ontwikkeling.
Vruchten worden voornamelijk verspreid door dieren (zoöchorie), de wind (anemochorie), water (hydrochorie) en bepaalde insecten, zoals mieren (myrmecochorie).
Vaak wordt in het normale spraakgebruik gesproken van zaden, waarbij het eigenlijk vruchtjes zijn. Zo wordt bij de composietenfamilie vaak gesproken van zaden (andijviezaad, zonnebloempitten enz.), terwijl het eenzadige vruchtjes zijn.
Vlezige vruchten
-
- bes (aardappel, tomaat, meloen, blauwe bosbes, druif, banaan)
- rozenbottel
- steenvrucht (kers, pruim, perzik, kokosnoot, braam, framboos)
- pitvrucht (appel, peer)
- komkommervrucht (komkommer)
- oranjevrucht (sinaasappel, citroen, sukade)
- samengestelde vrucht (ananas)
- Vlezige vruchten
-
 Steenvrucht: 'Reine Victoria' met gomgang in lengte doorgesneden.
Steenvrucht: 'Reine Victoria' met gomgang in lengte doorgesneden. -
 Besvrucht: tomaat in lengte doorgesneden
Besvrucht: tomaat in lengte doorgesneden -




Openspringende droge vruchten:
- kluisvrucht (openspringend, één zaad per vrucht)
- Driekluizig (wolfsmelk)
- Vijfkluizig (Geranium, springzaad)
- doosvrucht (openspringend, meer dan één zaad per vrucht)
- peul (vlinderbloemigen, zoals boon, erwt)
- kokervrucht (gewone dotterbloem, witte engbloem)
- hauw; minstens driemaal zo lang als breed (koolzaad, pinksterbloem)
- hauwtje; hoogstens driemaal zo lang als breed (judaspenning, herderstasje)
- Openspringende doosvrucht
- met kleppen (akkerviooltje, doornappel, klaverzuring, paardenkastanje)
- met tanden (dagkoekoeksbloem)
- met spleten (orchidee)
- met poriën (klaproos)
- met deksel (guichelheil)
- Openspringende droge vruchten
-

-
 Zaaddoos met aan de bovenkant geopende poriën van de klaproos.
Zaaddoos met aan de bovenkant geopende poriën van de klaproos. -
 Doosvrucht met haakjes van kleefkruid.
Doosvrucht met haakjes van kleefkruid.



Niet openspringende droge vruchten:
- dopvrucht (niet openspringend, één zaad per vrucht)
- enkele dopvrucht (scherpe boterbloem)
- noot vruchtwand is verhout (hazelnoot, eikel, beukennoot, linde)
- nootje dikwijls met vruchtpluis (composieten paardenbloem, klein kruiskruid)
- gevleugelde noot (es, iep)
- dubbele dopvrucht
- gevleugelde dopvrucht (esdoorn)
- veelvoudige dopvrucht (Aardbei)
- splitvrucht splitst zich bij rijpheid in afzonderlijke dopvruchten
- tweedelige splitvrucht (gewone berenklauw, fluitenkruid, zevenblad)
- gevleugelde splitvrucht (esdoorn)
- driedelige splitvrucht (Oost-Indische kers, moeraszoutgras)
- vierdelige splitvrucht (witte dovenetel, gewone smeerwortel)
- vijfdelige splitvrucht (gewone reigersbek)
- veeldelige splitvrucht (groot kaasjeskruid, muskuskaasjeskruid)
- tweedelige splitvrucht (gewone berenklauw, fluitenkruid, zevenblad)
- enkele dopvrucht (scherpe boterbloem)
- graanvrucht (grassen)
- sommige doosvruchten
- Niet openspringende droge vruchten
-
 Noten van hazelaar ras Géant de Halle.
Noten van hazelaar ras Géant de Halle. -
 Gevleugelde splitvruchten van rode noorse esdoorn.
Gevleugelde splitvruchten van rode noorse esdoorn. -
 Tarwekorrels, graanvrucht zonder kafjes
Tarwekorrels, graanvrucht zonder kafjes -
 Complexe vorm: aardbei in lengte doorgesneden
Complexe vorm: aardbei in lengte doorgesneden




Literatuur bewerken
Noten
Gebruikte literatuur
Noten Coniferen
Noten Gnetaceae
Noten Welwitschiaceae
Noten Ephedraceae
Literatuur Gnetales
| |||