Mechanoreceptor
Een mechanoreceptor is een zenuwreceptor die reageert op mechanische druk of vervorming. Mechanoreceptoren worden gestimuleerd door zenuwcellen die mechanische druk omzetten in elektrische signalen die bij dieren naar het centrale zenuwstelsel worden gestuurd. Ze zijn te vinden in sensorische organen , zoals de oren, de huid en de slagaders. Derhalve is er een grote verscheidenheid aan mechanoreceptoren die sterk variëren in structuur en functie.
Mechanoreceptoren van gewervelde dieren bewerken
De belangrijkste classificatie van mechanoreceptoren is gebaseerd op hun oorsprong. Eén deel van de mechanoreceptoren is ontstaan uit het epitheel en het andere deel uit de zenuwknopen.
Epithele mechanoreceptoren bewerken
Er wordt aangenomen dat epitheelreceptorcellen in de loop van de evolutie 2 tot 3 keer onafhankelijk zijn ontstaan en dat alle huidige sensorische cellen van dit type modificaties van dergelijke originele vormen vertegenwoordigen. In de huid voorkomende mechanoreceptoren reageren op mechanische stimuli die het gevolg zijn van fysieke prikkeling, waaronder druk en trillingen. Ze worden allemaal gestimuleerd door Aβ-vezels, behalve de mechanorecepterende vrije zenuwuiteinden, die worden gestimuleerd door Aδ-vezels. De in de huid voorkomende mechanoreceptoren kunnen worden ingedeeld op basis van het soort gevoel dat ze waarnemen, op basis van de snelheid van aanpassing en op basis van morfologie. Bovendien heeft elk een ander receptief veld.
Op tastzin bewerken
Verschillende gespecialiseerde lichaampjes en zenuwuiteinden gelegen in of vlak onder de huid, sturen signalen over de sensatie die ze waarnemen naar de hersenen. Vooral het sensorische projectiegebied is betrokken bij de verwerking van tastprikkels. De tast- en drukreceptoren bevinden zich overal in de huid, alleen zijn deze niet gelijkmatig verdeeld over ons lichaam. De vingers, voeten, lippen en tong hebben bijvoorbeeld een heleboel sensoren. Op de rug zitten er relatief weinig. Er bestaan verder fasische tastzintuigen die vooral gevoelig zijn voor veranderingen in druk en tonische tastzintuigen die signalen uitzenden zolang de prikkeling aanhoudt. De laatsten vindt men bijvoorbeeld rondom het hart waar zij de bloeddruk registreren.
Er is een indeling in zes soorten tactiele prikkels te maken, die alle zes ook gespecialiseerde receptoren in het epitheel kennen.
- De Slowly Adapting type 1 (SA1) mechanoreceptor, met het lichaampje van Merkel (ook bekend als Merkel-schijven) detecteert aanhoudende druk en ligt ten grondslag aan de waarneming van vorm en textuur via de huid. Tastzin zenuwuiteinden van dit type zijn langzaam adaptief, dat wil zeggen dat ze hun impulsen nog lang blijven afgeven naar de hersenen nadat de aanraking begon. Dit is ook nodig om zich bewust te blijven van de langdurige aanraking.[1] Ze hebben kleine receptieve velden en produceren langdurige reacties op statische stimulatie.
- De Slowly Adapting type 2 (SA2) mechanoreceptoren, met de lichaampjes van Ruffini (ook bekend als de bolvormig lichaampjes), detecteren spanning diep in de huid en fascia en reageren met het oprekken van de huid, maar is niet nauw verbonden met proprioceptieve of mechanoreceptieve rollen in perceptie.[2] Ze produceren ook aanhoudende reacties op statische stimulatie, maar hebben grote receptieve velden.
- De Rapidly Adapting (RA) of lichaampje van Meissner (ook bekend als de tastlichaampjes) ligt ten grondslag aan de perceptie van lichte aanraking zoals prikkels die onder meer verwerkt worden bij het uitoefenen van fijnmotorische taken door het organisme. De fijne tastzin kan getest worden door met een plukje watten zachtjes over de huid te strijken. De persoon dient de ogen te sluiten en de aanraking te bevestigen.[3][4] Het past zich snel aan aan veranderingen in textuur (trillingen rond 50 Hz). Ze hebben kleine receptieve velden en produceren voorbijgaande reacties op het begin en het einde van de stimulatie.
- Het lichaampje van Pacini of Vater-Paciniaanse lichaampje of lamellair lichaampje[5] in de huid en fascia detecteren snelle trillingen van ongeveer 200–300 Hz.[3][6] Hiermee kan een organisme bijvoorbeeld de trillingen van een aardbeving voelen. De mens bemerkt hiermee in de huid bijvoorbeeld de trillingen van de bas van een hard vibrerende muziekinstallatie. Deze lichaampjes bevinden zich in de handpalmen, voetzolen, geslachtsorganen en tepels, wat suggereert dat ze ook een belangrijke rol spelen bij de seksuele stimulatie en bij het zogen. Verder komen deze lichaampjes ook veel voor bij inwendige organen en in het bindweefsel rondom gewrichten en spieren, waar ze snelle veranderingen en vibraties registreren en betrokken zijn bij de proprioceptie. Ze produceren ook voorbijgaande reacties, maar hebben grote receptieve velden.
- Vrije zenuwuiteinden detecteren aanraking, druk, strekking, evenals de sensaties van kietelen en jeuk. Jeuksensaties worden veroorzaakt door stimulatie van vrije zenuwuiteinden door chemicaliën.[7] Ze bepalen mede de proprioceptie. Baroreceptoren registreren drukveranderingen in een orgaan en bestaan uit vrije zenuwuiteinden die zich vertakken binnen de elastische weefsels in de wanden van organen die kunnen uitrekken. Hoewel baroreceptoren onmiddellijk op drukveranderingen reageren vertonen ze ook een snelle adaptatie met als gevolg dat de output langs de sensorische vezels geleidelijk naar het normale niveau terugkeert.
- Haarzakje-receptoren, haarwortelplexi, voelen wanneer een haar van positie verandert. De gevoeligste mechanoreceptoren bij mensen zijn de haarcellen in het slakkenhuis van het binnenoor. Deze receptoren transduceren geluid naar de hersenen.[7] De zenuwuiteinden die om de haarzakjes gedraaid zitten registreren het bestrijken van de huid. Het gevoel dat deze peritrichale zenuwen veroorzaken is duidelijk op te wekken door tegen de natuurlijke stand van de hoofdharen in te strijken.
-
 Lichaampje van Merkel
Lichaampje van Merkel -
 Lichaampje van Ruffini
Lichaampje van Ruffini -
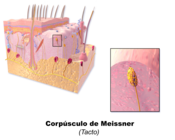 Lichaampje van Meissner
Lichaampje van Meissner -
 Lichaampje van Pacini
Lichaampje van Pacini -
 Vrije zenuwuiteind
Vrije zenuwuiteind -
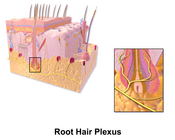 Haarwortelplexus
Haarwortelplexus






Op aanpassingssnelheid bewerken
Epithele mechanoreceptoren kunnen ook in categorieën worden onderverdeeld op basis van hun aanpassingssnelheid. Wanneer een mechanoreceptor een stimulus ontvangt, begint deze impulsen of actiepotentialen af te vuren met een verhoogde frequentie (hoe sterker de stimulus, hoe hoger de frequentie). De cel zal zich echter snel "aanpassen" aan een constante of statische stimulus en de impulsen zullen afnemen tot een normaal tempo. Receptoren die zich snel aanpassen (dat wil zeggen snel terugkeren naar een normale impulsfrequentie) worden "fasische" genoemd. De receptoren die langzaam terugkeren naar hun normale impulsfrequentie worden tonische genoemd. Fasische mechanoreceptoren zijn nuttig bij het waarnemen van zaken als textuur of trillingen, terwijl tonische receptoren onder meer nuttig zijn voor het waarnemen van temperatuur en voor de proprioceptie.
- Langzaam aanpassend: Langzaam aanpassende mechanoreceptoren omvatten lichaampje van Merkel en lichaampje van Ruffini en enkele vrije zenuwuiteinden.
- Traag aanpassende mechanoreceptoren van type I hebben meerdere lichaampjes van Merkel.
- Traag aanpassende type II mechanoreceptoren hebben enkele lichaampjes van Ruffini.
- Intermediaire aanpasing: Sommige vrije zenuwuiteinden zijn intermediair aanpassend.
- Snel aanpassend: Snel aanpassende mechanoreceptoren omvatten het lichaampje van Meissner, lichaampje van Pacini, haarzakjesreceptoren en enkele vrije zenuwuiteinden.
- Snel aanpassende mechanoreceptoren van type I hebben meerdere lichaampjes van Meissner
- Snel aanpassende type II mechanoreceptoren (meestal Pacini genoemd) hebben enkele lichaampjes van Pacini.
Via receptief veld bewerken
Epithele mechanoreceptoren met kleine, nauwkeurige receptieve velden worden aangetroffen in gebieden die een nauwkeurige tast vereisen (bijvoorbeeld op de vingertoppen). In de vingertoppen en lippen is de innervatiedichtheid van langzaam aanpassende type I en snel aanpassende type I mechanoreceptoren groot. Deze twee soorten mechanoreceptoren hebben kleine discrete receptieve velden en er wordt aangenomen dat ze ten grondslag liggen aan het meeste laagdrempelige gebruik van de vingers bij het beoordelen van textuur, wrijving en trillingen. Mechanoreceptoren die worden aangetroffen in delen van het lichaam met minder tastscherpte hebben doorgaans grotere receptieve velden.
Rekmechanoreceptoren bewerken

Rekreceptoren zijn mechanoreceptoren, die reageren op uitzetting van verschillende organen en rek van spieren en zijn neurologisch verbonden met de medulla in de hersenstam via afferente zenuwvezels. Voorbeelden hiervan zijn rekreceptoren in de arm- en beenspieren en pezen, in het hart, in de dikke darmwand en in de longen.
Rekreceptoren worden ook aangetroffen rond de halsslagader, waar ze de bloeddruk controleren en de afgifte van antidiuretisch hormoon (Antidiuretisch hormoon) uit de hypofyseachterkwab stimuleren.
Rekreceptoren zijn onder meer:
- Golgi-peeslichaam
- Spierspoeltjes, zijn vooral te vinden tussen de spiervezels van spieren die verantwoordelijk zijn voor de lichaamshouding (Proprioceptie).
- Pulmonale rekreceptoren, worden in de longen aangetroffen
- Chordotonaal orgaan, bij insecten. Interne rekreceptoren bij de gewrichten kunnen zowel extero- als proprioceptieve functies hebben
De Golgi-peeslichamen geven signalen door aan de hersenen over de rek in pezen. Daardoor wordt de maximale uitslag van een pees onbewust gecontroleerd en overbelasting voorkomen. Ze komen ook voor in alle andere bindweefselvliezen.
Spierspoeltjes registreren rek of verkorting van spieren en zijn bij de proprioceptie betrokken.
Lamellaire lichaampjes bewerken
Lamellaire lichaampjes of lichaampjes van Pacini of lichaampjes van Vater-Pacini zijn vervormings- of drukreceptoren die in de huid en ook in verschillende inwendige organen zitten.[8] Elk lichaampje is verbonden met een sensorisch neuron. Vanwege de relatief grote omvang ervan kan een enkel lamellair lichaampje worden geïsoleerd en de eigenschappen ervan worden bestudeerd. Mechanische druk met variërende sterkte en frequentie kan met een drukpen op het lichaampje worden uitgeoefend en de resulterende elektrische activiteit kan worden gedetecteerd door elektroden die aan het preparaat zijn bevestigd.
Door het lichaampje te vervormen ontstaat er een receptorpotentiaal in de sensorische neuron. Dit is een graduele respons: hoe groter de vervorming, hoe groter het receptorpotentiaal. Als het receptorpotentiaal de drempel bereikt, wordt een salvo van actiepotentialen (zenuwimpulsen) geactiveerd bij de eerste insnoering van Ranvier van het axon.
Zodra de drempel is bereikt, wordt de omvang van de stimulus omgezet in de frequentie van de impulsen die in het neuron worden gegenereerd. Dus hoe groter of sneller de vervorming van een enkel lichaampje, des te hoger de frequentie van zenuwimpulsen die in het neuron ervan worden gegenereerd.
De optimale gevoeligheid van een lamellair lichaampje is 250 Hz. Het frequentiebereik dat op de vingertoppen wordt gegenereerd door structuren kleiner dan 200 micrometers.[9]
Ligamente mechanoreceptoren bewerken
Er zijn vier soorten mechanoreceptoren ingebed in ligamenten. Omdat al deze soorten mechanoreceptoren ingebed zijn in myeline, kunnen ze snel sensorische informatie over gewrichtsposities doorgeven aan het centrale zenuwstelsel.[10]
- Type I: (klein) Lage drempel, langzaam aanpassend in zowel statische als dynamische omgevingen
- Type II: (gemiddeld) Lage drempel, past zich snel aan in dynamische omgevingen
- Type III: (groot) Hoge drempel, langzaam aanpassend in dynamische instellingen
- Type IV: (zeer kleine) Pijnreceptoren of Nociceptoren met een hoge drempel die letsel communiceren
Er wordt aangenomen dat met name type II- en type III-mechanoreceptoren verband houden met iemands gevoel van proprioceptie.
Mechanoreceptoren van ongewervelden bewerken

Mechanoreceptoren voor insecten en geleedpotigen zijn onder meer:
- Campaniforme sensilla: kleine koepels in het exoskelet die over het hele lichaam van het insect zijn verdeeld. Er wordt aangenomen dat deze cellen mechanische belasting detecteren als weerstand tegen spiercontractie, vergelijkbaar met de Golgi-peeslichamen van zoogdieren.
- Haarplaten: sensorische neuronen die haren innerveren die zich in de plooien van insectengewrichten bevinden. Deze haren worden afgebogen wanneer een lichaamssegment beweegt ten opzichte van een aangrenzend segment. Ze hebben een proprioceptieve functie en er wordt aangenomen dat ze fungeren als limietdetectoren die het extreme bewegingsbereik van elk gewricht coderen.
- Chordotonale organen: Interne rekreceptoren bij de gewrichten kunnen zowel extero- als proprioceptieve functies hebben. De neuronen in het cordotonale orgaan van Drosophila melanogaster kunnen worden georganiseerd in knuppel-, klauw- en haakneuronen. Er wordt aangenomen dat knuppelneuronen trillingssignalen coderen, terwijl klauw- en haakneuronen kunnen worden onderverdeeld in extensie- en flexiepopulaties die respectievelijk de gewrichtshoek en beweging coderen.
- Spleet sensilla: Spleten in het exoskelet die fysieke vervorming van het exoskelet van het dier detecteren, hebben een proprioceptieve functie
- Borstelhaar sensilla: Borstelneuronen zijn mechanoreceptoren die haren over het hele lichaam innerveren. Elk neuron verlengt een dendritisch proces om een enkele haar te innerveren en projecteert zijn axon naar het ventrale zenuwkoord. Er wordt aangenomen dat deze neuronen de aanrakingssensatie bemiddelen door te reageren op fysieke afbuigingen van het haar. In overeenstemming met het feit dat veel insecten haren van verschillende grootte vertonen, gewoonlijk macrochaeten (dikkere langere haren) en microchaeten (dunnere, kortere haren) genoemd, suggereren eerdere onderzoeken dat borstelneuronen van deze verschillende haren verschillende prikkeleigenschappen kunnen hebben, zoals rustmembraan potentieel en prikkeldrempel.
-
 Campaniforme sensillum
Campaniforme sensillum -
 Haarplaat
Haarplaat -
 Diagram van de primaire onderdelen van een chordotonaal orgaan: het scolopidium
Diagram van de primaire onderdelen van een chordotonaal orgaan: het scolopidium -
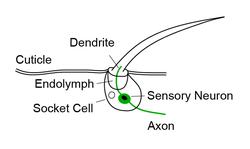 Borstelhaar sensillum
Borstelhaar sensillum




Mechanoreceptoren bij planten bewerken
Mechanoreceptoren zijn ook aanwezig in plantencellen waar ze een belangrijke rol spelen bij de normale groei, ontwikkeling en het waarnemen van hun omgeving.[11] Mechanoreceptoren op de voelharen helpen de venusvliegenvanger (Dionaea muscipula) bij het vangen van prooien.[12][13]
Referenties bewerken
- ↑ (en) Johnson KO, Hsiao SS (1992). Neural mechanisms of tactual form and texture perception. Annual Review of Neuroscience 15: 227–50. PMID 1575442. DOI: 10.1146/annurev.ne.15.030192.001303.
- ↑ (en) Torebjörk HE, Ochoa JL (December 1980). Specific sensations evoked by activity in single identified sensory units in man. Acta Physiologica Scandinavica 110 (4): 445–7. PMID 7234450. DOI: 10.1111/j.1748-1716.1980.tb06695.x.
- ↑ a b (en) Talbot WH, Darian-Smith I, Kornhuber HH, Mountcastle VB (March 1968). The sense of flutter-vibration: comparison of the human capacity with response patterns of mechanoreceptive afferents from the monkey hand. Journal of Neurophysiology 31 (2): 301–34. PMID 4972033. DOI: 10.1152/jn.1968.31.2.301.
- ↑ (en) Johansson RS, Westling G (1987). Signals in tactile afferents from the fingers eliciting adaptive motor responses during precision grip. Experimental Brain Research 66 (1): 141–54. PMID 3582528. DOI: 10.1007/bf00236210.
- ↑ (en) Biswas A, Manivannan M, Srinivasan MA (2015). Multiscale layered biomechanical model of the pacinian corpuscle. IEEE Transactions on Haptics 8 (1): 31–42. PMID 25398182. DOI: 10.1109/TOH.2014.2369416.
- ↑ (en) Biswas A, Manivannan M, Srinivasan MA (2015). Vibrotactile sensitivity threshold: nonlinear stochastic mechanotransduction model of the Pacinian Corpuscle. IEEE Transactions on Haptics 8 (1): 102–13. PMID 25398183. DOI: 10.1109/TOH.2014.2369422.
- ↑ a b (en) Tortora GJ (2019), Principles of anatomy and physiology. John Wiley & Sons Australia, Limited. ISBN 978-0-7303-5500-7.
- ↑ Biswas A (2015). Characterization and Modeling of Vibrotactile Sensitivity Threshold of Human Finger Pad and the Pacinian Corpuscle (PhD). Indian Institute of Technology Madras, Tamil Nadu, India. doi:10.13140/RG.2.2.18103.11687.
- ↑ (en) Scheibert J, Leurent S, Prevost A, Debrégeas G (March 2009). The role of fingerprints in the coding of tactile information probed with a biomimetic sensor. Science 323 (5920): 1503–6. PMID 19179493. DOI: 10.1126/science.1166467.
- ↑ (en) Michelson JD, Hutchins C (March 1995). Mechanoreceptors in human ankle ligaments. The Journal of Bone and Joint Surgery. British Volume 77 (2): 219–24. PMID 7706334. DOI: 10.1302/0301-620X.77B2.7706334.
- ↑ (en) Monshausen GB, Haswell ES (November 2013). A force of nature: molecular mechanisms of mechanoperception in plants. Journal of Experimental Botany 64 (15): 4663–80. PMID 23913953. PMC 3817949. DOI: 10.1093/jxb/ert204.
- ↑ (en) Chamovitz D (2012), What a plant knows : a field guide to the senses, 1st. Scientific American/Farrar, Straus and Giroux, New York. ISBN 9780374533885.
- ↑ (en) Volkov AG, Forde-Tuckett V, Volkova MI, Markin VS (10 februari 2014). Morphing structures of the Dionaea muscipula Ellis during the trap opening and closing. Plant Signaling & Behavior 9 (2): e27793. PMID 24618927. PMC 4091236. DOI: 10.4161/psb.27793.