Gigantopithecus
Gigantopithecus[1][2] is een geslacht van uitgestorven primaten uit de groep van de mensapen (Hominidae).
| Gigantopithecus Status: Uitgestorven, als fossiel bekend Fossiel voorkomen: Midden-Pleistoceen | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|

| |||||||||||||||||||
| Taxonomische indeling | |||||||||||||||||||
| |||||||||||||||||||
| Geslacht | |||||||||||||||||||
| Gigantopithecus † Von Koenigswald, 1935 | |||||||||||||||||||
| Typesoort | |||||||||||||||||||
| Gigantopithecus blacki | |||||||||||||||||||
| |||||||||||||||||||
| Afbeeldingen op | |||||||||||||||||||
| Gigantopithecus op | |||||||||||||||||||
| |||||||||||||||||||
Het geslacht kreeg in 1935 zijn naam die 'reusachtige aap' betekent, toen de typesoort Gigantopithecus blacki benoemd werd door Ralph von Koenigswald. In Hongkong kocht hij dat jaar in een groothandel voor medicijnen een aantal 'drakentanden' waaronder een enorme kies. Sinds de jaren vijftig werden in grotten in Zuid-China nog een kleine tweeduizend losse tanden gevonden alsmede vier onderkaken. De fossielen stammen uit het Midden-Pleistoceen en hebben een ouderdom van twee miljoen tot driehonderdduizend jaar. Op basis van een tand en een onderkaak uit het Boven-Mioceen, acht tot zeven miljoen jaar oud, van Noord-India en Pakistan werd in 1969 een tweede soort benoemd: Gigantopithecus bilaspurensis die ook wel Gigantopithecus giganteus genoemd werd. Tegenwoordig wordt dit als het aparte geslacht Indopithecus beschouwd.
Omdat de vondsten zo beperkt zijn, is weinig met zekerheid bekend over de bouw van Gigantopithecus. Eerst werd een monsterlijke omvang verondersteld met een geschatte hoogte van 3,7 meter. Later werd dat bijgesteld tot rond de twee meter bij een gewicht van ongeveer tweehonderdvijftig kilogram. Dat maakt hem nog steeds de grootste primaat die bekend is. Zijn hoektanden zijn relatief klein. De kiezen zijn extreem groot met zeer dik glazuur. Ze kunnen gebruikt zijn om harde noten te kraken of taaie wortels te kauwen. De krassen op het glazuur en de samenstelling ervan wijzen er echter op dat voornamelijk vruchten gegeten werden. Dat past ook beter bij zijn leefgebied, dichte subtropische wouden. Wellicht dat Gigantopithecus uitstierf toen de bossen in omvang verminderden. Hij kan ook uitgeroeid zijn door de eerste mensachtige die zijn gebied bereikte, Homo erectus.
Gigantopithecus is een lid van de Ponginae. Zijn nauwste levende verwanten zijn de orang-oetans. Onder de fossiele soorten is Sivapithecus nauw verwant. Dat werd al onderkend door Von Koenigswald. Er is echter een periode geweest dat veel onderzoekers dachten dat het om een tweevoetige mensachtige ging, zelfs een voorouder van de moderne mens. Dat heeft veel bijgedragen aan het mysterieuze karakter van de soort. De kryptozoölogie sloot hierbij aan door te veronderstellen dat Gigantopithecus verantwoordelijk was voor de legenden over de verschrikkelijke sneeuwman en bigfoot. Biologen achten dit zeer onwaarschijnlijk.
Vondst en naamgeving bewerken
Betekenis van de naam bewerken

Gigantopithecus is een neologisme. De naam van het geslacht is afgeleid van de Griekse woorden πίθηκος (oud-Grieks uitgesproken als píthēkos): 'aap' en γίγας, gígas 'reus'.
Het epitheton van het type, Gigantopithecus blacki, eert de Canadese arts en paleoantropoloog Davidson Black, die stierf in 1934, 'wiens fundamentele werkzaamheden aangaande Sinanthropus hem een blijvende herinnering zal verzekeren en wie het helaas niet meer gegund was zijn werk te voltooien'. Gigantopithecus blacki betekent dus 'Black's gigantische aap'.
Toen Gigantopithecus pas benoemd was, dachten veel geleerden dat hij nauw aan de mens verwant was. Sommigen vonden de naam daarom ongepast. In 1946 suggereerde Franz Weidenreich de naam Gigantanthropus, wat in 1950 weer door Hans Weinert gewijzigd werd in Giganthropus. Beide namen zijn jongere synoniemen van Gigantopithecus, welke prioriteit heeft en aldus de geldige naam blijft.
Gigantopithecus wordt benoemd bewerken
Het genoholotype van het geslacht en tegelijkertijd het holotype van de soort Gigantopithecus blacki is een derde rechterkies (M3) uit de onderkaak. De vondst daarvan is een van de befaamdste verhalen uit de paleoantropologie en een van de weinige met een verband met Nederland. In 1930 was de Duitse paleontoloog Gustav Heinrich Ralph von Koenigswald in Nederlands-Indië gaan werken voor de Dienst van Mijnbouw van Nederlands-Indië. Daarvoor was hij bij het Paläontologisches Museum München assistent geweest van professor Max Schlosser bij het ordenen van de collectie van Karl Albert Haberer die tijdens een reis tussen 1899 en 1901 een grote verzameling fossielen in China had gekocht van apotheken in Peking en Shanghai. Het was al sinds het midden van de negentiende eeuw bekend dat in apotheken voor traditionele Chinese geneeskunde interessant materiaal te vinden was. Destijds golden fossielen in China als zogenaamde 'drakenbotten' die tot poeder werden gemalen omdat er een helende werking aan toegeschreven werd. Von Koenigswald kon nu zulke apotheken op Java zelf bezoeken en kocht wat fossielen. In 1935 bezocht hij eerst een apotheek in Manilla en toen een groothandel in medicijnen te Hongkong en nam de gelegenheid te baat om daar wat ruimer in te slaan, twee kuub aan fossielen kopend. In een partij van een vijftigtal fossiele tanden van orang-oetans viel hem daar een enorme kies op. Een andere grote derde kies, uit de bovenkaak, werd iets later in Zuid-China verworven maar kon in een addendum bij het benoemende artikel nog vermeld worden. Het is niet bekend waar de desbetreffende groothandel in Hong Kong precies de kiezen vandaan had.

In hetzelfde jaar publiceerde Von Koenigswald de formele eerste beschrijving van het geslacht Gigantopithecus, dat hij dus tegelijkertijd benoemde, die echter slechts een paar regels lang was. Het artikel werd in juli 1935 geschreven en op 28 september 1935 door professor Louis Rutten voorgedragen op een vergadering van de Koninklijke Akademie van Wetenschappen. Het holotype maakte deel uit van de collectie van de Universiteit van Utrecht, samen met een zevental verdere tanden. Ze bleven wel eigendom van Von Koenigswald. Von Koenigswald ging na zijn verblijf in een interneringskamp tijdens de Tweede Wereldoorlog in Amerika en Nederland werken. Pas na de oorlog kon hij in 1952 de soort meer in detail beschrijven. Na zijn dood werd zijn collectie ondergebracht bij het Senckenberg Gesellschaft für Naturforschung.

De kies viel bij Von Koenigswald op omdat deze veel groter was dan alle orang-oetantanden en zich duidelijk van ze onderscheidde door een veel grover kroonreliëf. De tand was ook groter dan die van een gorilla, maar was flink versleten. Zijn grootste lengte is tweeëntwintig millimeter en zijn grootste breedte is achttien millimeter. De kies mist niet alleen volledig het sterk gerimpelde email dat typisch is voor apen, merkt Von Koenigswald op in de eerste beschrijving, hij wordt ook gekenmerkt door een vreemde overontwikkeling van secundaire knobbels, waardoor het een enigszins vreemde verschijning kreeg voor een primaattand. Als men echter een diagram van de tand tekent, blijken het dezelfde secundaire bultjes te zijn die ook in de orang-oetan kunnen voorkomen. Zo heeft de kies een typisch tuberculum accessorium mediale internum, dus een extra knobbeltje aan de tongzijde en de kant van de voorliggende kies.
In 1938 kocht Von Koenigswald op Java een bovenste hoektand. In 1939 verwierf hij in Hong Kong een onderste derde kies die niet versleten was. Tegen 1939 had Von Koenigswald nog drie individuele Gigantopithecus-tanden in Chinese apotheken ontdekt: een bovenste eerste voortand, nog een bovenste hoektand en twee onderste vierde premolaren. Al die tanden konden alleen op grond van hun grootte gedetermineerd worden.
In 1935 begon, zoals Von Koenigswald nog meekreeg, het Chinese Cenozoic Laboratory of the Geological Survey of China grotten te onderzoeken in het zuiden van China, in Guangdong en Guangxi, waarbij fossielen van de Stegodon-Ailuropoda-fauna gevonden werden. Dit maakte het uiteindelijk mogelijk de tanden te dateren want de fossielen die de apotheken verwierven maakten duidelijk deel uit van die fauna. Zowel de verhandelde als de gevonden fossielen waren ook aangevreten door stekelvarkens. De aanwezigheid van de fossielen is wel verklaard door aan te nemen dat in het Pleistoceen stekelvarkens, van het geslacht Hystrix, ze naar hun holen in een grot sleepten om ze aan te knagen teneinde hun kalkbehoefte te dekken voor de aanmaak van hun stekels. De tandkronen, waarvan de wortels verdwenen zijn, zouden overgebleven zijn omdat ze te hard waren. Fossiele orang-oetans zijn alleen van zulke tanden bekend.
Verdere vondsten bewerken

In 1955 werd de Chinese onderzoeker Pei Wenchung door het Institute of Vertebrate Paleontology and Paleoanthropology uitgezonden om de exacte herkomst van de tanden vast te stellen. Zevenenveertig tanden werden in beslag genomen bij lokale fossielenhandelaren. Die verklapten de locatie van zo'n driehonderd grotten in de karstlandschappen van Zuid-China. In achtentachtig daarvan lokaliseerde Pei inderdaad zoogdierfossielen. In 1956 vond Pei in de Heidong, de 'Duistere Grot' op de berg Niusui, drie tanden, de eerste die in situ door paleontologen werden opgegraven. In de zomer van dat jaar groef boer Qin Xiuhuai in Gunagxi de bodem van de Xiaoyangrot in Liucheng (柳城) uit als meststof. Fossielen die hij aantrof, probeerde hij te verkopen als drakenbeenderen maar merkte dat dit intussen verboden was. Bankdirecteur Wei Yaoshe zag dat er een onderkaak tussen zat en overtuigde Qin die aan de autoriteiten af te staan. Thuisgekomen doorzocht Qin met zijn kinderen het veld waar de mest was uitgereden en vond nog een aantal tanden die uit de kaak waren gevallen. Dit was de eerste complete onderkaak die van Gigantopithecus gevonden werd. Het is specimen PA77, ook wel als Mandibula I aangeduid, en is vermoedelijk van een volwassen vrouw. Tussen 1957 en 1963 groef Pei de grot verder uit en vond ruim duizend Gigantopithecus-tanden en nog twee gepaarde onderkaken: Mandibula II ofwel PA78 is van een jongvolwassen individu; Mandibula III ofwel PA83 is van een volwassen man en de grootste onderkaak die bekend is. Deze 'gigantopithecusgrot' werd in 1963 door het bestuur van Guangxi aangemerkt als officieel paleontologisch erfgoed.

In de jaren zestig en zeventig werden nog meer groeven in grotten geopend. In 1965 werden in een grot op de Bulaliberg twaalf tanden gevonden. Bij boeren in Hubei werden nog eens tweehonderd tanden in beslag genomen. In 1970 zond het IVPP een team naar Hubei om de Longgudong uit te graven, de 'drakenbottengrot', wat nog eens drieëndertig tanden opleverde. Tussen 1998 en 2000 werd de grot opnieuw onderzocht waarbij achtentwintig tanden geborgen werden. In 1973 werd een tand gevonden in een grot op de Nomoberg in het Bamadistrict. In 1974 werden zes tanden gemeld uit de Xiniugrot, de 'neushoorngrot'. Ondanks het vele materiaal bleef de kennis beperkt tot tanden en onderkaken. Van de eigenlijke schedel en het postcraniaal skelet was niets aangetroffen wat bijdroeg aan de mysterieuze sfeer die rond Gigantopithecus bleef hangen.
in de jaren zestig werden in grotten in Noord-Vietnam tanden gevonden die toegewezen werden aan Gigantopithecus. Achteraf bleken ze van de uitgestorven orang-oetansoort Pongo weidenreichi te zijn. In 2022 werden opnieuw resten aan Gigantopithecus toegewezen die misschien de jongste bekende vondsten vertegenwoordigen.
De belangstelling voor Gigantopithecus zakte na de jaren zeventig in China wat in. In 1964 had een team van het IVPP wat fossielen verworven van apotheken in Bijie in Guizhou. Pas in de jaren tachtig bleek het Xu Chunhua dat er een bovenste vierde promolaar van Gigantopithecus tussen zat. Hierop werd de Ba'eryangrot bij het dorp Guankou onderzocht en in 2006 kon de vondst van nog eens vijf tanden gemeld worden. In 2004 ontdekte een team van de bioloog professor Pan Wenshi in de Juyuangrot op de Boyueberg bij toeval fossielen. Paleontoloog Qiu Zhanqian herkende er tanden van Gigantopithecus onder. Dat leidde tot een nieuwe golf aan ontdekkingen in de regio Chongzuo. In de grotten van Baikong, Yanliang, Sanhe, Queque, Shuangtan en Hejiang werden vierhonderd tanden opgegraven. De Yanlinaggrot leverde tegen 2016 de vierde onderkaak op die van de soort bekend is: specimen PA1601, waarbij al bij het leven een tweede kies verloren ging. Daarnaast werden er in het Bubingbasin in de Mohuigrot zestien tanden gevonden en in de Chuifengrot tweeënnegentig tanden. Verder werd er in 2006 een onderste kies gevonden in de Xinchonggrot op Hainan. Het eiland was indertijd verbonden met het vasteland. Dit zou het meest zuidelijke voorkomen van Gigantopithecus vertegenwoordigen.
Gigantopithecus bilaspurensis/giganteus bewerken
In 1969 werd er een tweede soort benoemd: Gigantopithecus bilaspurensis op basis van een onderkaak gevonden bij Haritalyangar in India. Eerder was door Pilgrim een grote kies benoemd als Dryopithecus giganteus. In 1937 werd die echter toegewezen aan een groot exemplaar van Sivapithecus indicus. In 1950 maakte Von Koenigswald daar een eigen geslacht van: Indopithecus. In 1979 werd onderkend dat G. bilaspurensis identiek was aan D. giganteus. Hierna sprak men wel van een Gigantopithecus giganteus. Sinds 2003 is het echter gebruikelijk geworden toch de naam Indopithecus te gebruiken.
Voorkomen en ouderdom bewerken
_Fundort_Nariokotome,_Kenia,_Rekonstruktion_im_Neanderthal_Museum.jpg)
Het gebied waar Gigantopithecus gevonden is, beperkt zich tot het zuiden van China maar beslaat desalniettemin een geweldige oppervlakte van zeshonderdduizend vierkante kilometer. De meest noordelijke vindplaatsen, Longgudong en Longgupo, liggen net ten zuiden van de Yangtze. De meest zuidelijke zekere vindplaats ligt op Hainan. Onzeker zijn vondsten uit Thailand zoals resten die in 2009 bij Had Pu Dai gemeld zijn. China was in het Pleistoceen naar het zuiden toe verbonden met een enorm gebied dat, afhankelijk van de hoogte van de zeespiegel, zich tot Java uitstrekte. Dat vormde alles een potentiële habitat. Dit wordt bevestigd door de melding uit 2016 dat in Semono in 2014 twee stukken onderkaak gevonden zijn met tanden van een gigantische mensaap. De tanden verschillen in detail van die uit China maar benaderen onder alle mensapen die van Gigantopithecus het dichtst. Het gaat of om een zuidelijke populatie van G. blacki of om een nauw verwante soort. Hoewel de populatiedichtheid van soorten met grote individuen meestal vrij laag ligt, kan de totale populatiegrootte in het hele gebied vele honderdduizenden hebben bedragen.
De oudste vondsten zijn uit de Longgugrot en mogelijk 2,15 miljoen jaar oud, dus uit het vroege Pleistoceen. De jongste vondsten zijn uit de Heigrot en kunnen zo jong zijn als 310 000 jaar. Ze stammen dus uit het middelste Pleistoceen. Uit het late Pleistoceen zijn geen vondsten bekend. Natuurlijk moet de populatie ouder zijn dan 2,15 miljoen jaar en eerder zal de soort zich van een andere tak hebben afgesplitst. Het is echter niet bekend welke dat is. Ook is onduidelijk wanneer en waarom de soort is uitgestorven. Evolutionaire veranderingen zoals een vergroting van de tanden suggereren dat het voedselaanbod zich wijzigde. Wellicht dat dit uiteindelijk tot het uitsterven leidde. Dit wordt echter niet ondersteund door onderzoek naar de flora. De rest van de fauna overleefde wellicht wel tot in het late Pleistoceen, hoewel een studie uit 2017 concludeerde dat die vierhonderdduizend jaar geleden verdween. Het is ook geopperd dat het arriveren van de mensensoort Homo erectus het einde betekende voor Gigantopithecus door overbejaging. De grote mensaap moet een aantrekkelijke prooi geweest zijn voor de kleinere: hij verschafte in een keer een ruime hoeveelheid vlees en was wellicht eenvoudig met speren te doden omdat hij traag was en niet goed kon klimmen. Ook hiervoor bestaat echter weinig direct bewijs. Homo erectus lijkt weinig te zijn voorgekomen in de wouden van Zuid-China. De twee miljoen jaar oude 'Wushanmens' bleek later een basale mensaap te zijn geweest.
Beschrijving bewerken
Grootte bewerken

Gigantopithecus is een geliefd onderwerp geworden in populair-wetenschappelijke boeken en wordt daar voorgesteld als een waar monster van ruim drie meter hoog met een gewicht van ruim een halve ton. Deze primaat wordt aldus gerekend tot de megafauna, de dierengemeenschap tussen een miljoen jaar en honderdduizend jaar geleden waarvan de leden een stuk groter waren dan de huidige dieren. Over grootte en gewicht kunnen naar moderne inzichten geen definitieve uitspraken worden gedaan, omdat tot nu toe alleen kaakelementen en tanden zijn gevonden. Deze zijn echter aanzienlijk groter dan hun tegenhangers in zowel levende als uitgestorven apen. Het is zeer waarschijnlijk dat de soort het grootste bekende lid van de Hominoidea is of van de primaten in het algemeen.
De grote vermeende lichaamsomvang zoals vermeld in populaire werken had echter zijn bron in oudere wetenschappelijke literatuur. Toen alleen een kies bekend was die leek op menselijke molaren, leidde dat tot speculaties dat er een soort oermens had rondgelopen met proporties die leken op Homo sapiens en een absolute lengte die tweemaal hoger lag. Weidenreich schatte in 1945 dat het gewicht tweemaal dat van een gorilla bedroeg. Mannelijke gorilla's wegen gemiddeld 169 kilogram. Zelfs toen duidelijk werd dat de proporties die van een typische basale mensaap waren bleven de schattingen hoog: voor het gewicht tussen de 225 en 300 kilogram. In 1957 stelde Pei nog dat een hoogte van 3,7 meter bereikt kon worden en in 1970 dacht Simons aan een lengte van 2,7 meter.
Geleidelijk kwam men tot meer realistische schattingen met aanzienlijk kleinere maten. In 1979 baseerde Alfred E. Johnson zich op de correlatie tussen de gorilla-onderkaak en de pijpbeenderen van gorillaledematen. De lengte van de pijpbeenderen van Gigantopithecus werd door hem geëxtrapoleerd op twintig tot vijfentwintig procent hoger dan die van gorilla's: 544 millimeter voor het dijbeen en 627 millimeter voor het opperarmbeen. Met vergelijkbare lichaamsverhoudingen zou dit overeenkomen met een hoogte van ongeveer honderdtachtig centimeter. Johnson dacht dat Gigantopithecus robuustere beenderen zou hebben gehad dan gorilla's. In 2015 analyseerde Zhang de beschikbare gegevens nog eens. Hij begon met opnieuw de kiezen als uitgangspunt te nemen. Extrapoleren uit de maten van de eerste kies volgens de methode Gingerich, die een formule had opgesteld gebaseerd op de correlaties bij moderne primaten, leverde een gewicht van tweehonderdvier kilogram op. De alternatieve methode Conroy kwam uit op tweehonderdtachtig kilogram. Zhang wees erop dat beide methoden vermoedelijk het gewicht overschatten omdat Gigantopithecus duidelijk megadont is, dus relatief grote kiezen bezit ten opzichte van het skelet als geheel. Desalniettemin achtte hij het waarschijnlijk dat het gewicht hoger lag dan bij gorilla's. Aangezien de zwaarste gorilla's ruim tweehonderd kilogram wegen, vond hij een massa van tussen de tweehonderd en driehonderd kilogram voor Gigantopithecus een redelijke inschatting. Zo'n gewicht verminderde vermoedelijk het vermogen tot klimmen.
Onderscheidende kenmerken bewerken
Zhang gaf in 2017 een aantal onderscheidende kenmerken aan. De gepaarde onderkaken zijn massief gebouwd met een sterk van achteren verstevigde symfyse en hoge en robuuste hoofdlichamen. Het onderste deel van het gezicht is kort en diep, afgaande op de onderkaakslengte. De diastemata in de onderkaak, de hiaten tussen de hoektanden en de derde premolaren waar bij verwanten de bovenste hoektanden in rusten, zijn klein of geheel afwezig. De voortanden zijn smal en verticaal in de kaak geplaatst met lage pinvormige kronen. De bovenste en onderste hoektanden zijn laag maar breed. De onderste derde premolaar heeft twee knobbels en een ten opzichte van verwanten gewijzigd facet aan de lipzijde van de kant van de onderste hoektand voor het contact met de bovenste hoektand. De onderste vierde premolaar is groot en benadert een kies in bouw. De onderste kiezen zijn lang en nauw met een sterke insnoering van lipzijde naar tongzijde, een vernauwing richting achterste tandenrij en een klein hypoconulide, een knobbeltje richting achterste tandenrij naast de hypocoon, de knobbel aan de lipzijde. De bovenste kiezen hebben slechts een beperkte insnoering en vernauwing. De bovenste en onderste kiezen hebben hoge kronen met lage gezwollen blokvormige knobbels die het hele kauwvlak vullen. Hun verbindende kammen zijn laag, afgerond en slecht ontwikkeld. De uithollingen op het kauwvlak zijn ondiep en weinig ontwikkeld. Tussen de knobbels liggen diepe groeven. Het email is zo geplooid dat secundaire knobbeltjes gevormd worden waardoor het vlak de indruk geeft van een patroon van meervoudige hoofdknobbels. De kiezen hebben dik tandemail dat aan de onderzijde slechts door korte hoorns van dentine ingekeept wordt. Zowel bij de bovenkaak als de onderkaak is de derde kies kleiner dan de tweede kies. Volgens Zhang zouden deze kenmerken kunnen gelden als autapomorfieën, unieke afgeleide eigenschappen.
Skelet bewerken
Onderkaak bewerken

De vier gevonden mandibulae geven een goed beeld van de bouw van de voorste onderkaak. De achterste opgaande takken zijn niet bewaard gebleven. Alleen het corpus mandibulae is aldus bekend. Dit corpus is hoog en het hoogst bij de mannelijke Mandibula III, veertig procent hoger dan bij de vrouwelijke Mandibula I. Naar achteren neemt de hoogte toe, de grootste verticale lengte bereikend onder de derde kies. De mandibula bestaat uit twee gepaarde onderkaken die bij Anthropoidea vooraan verbonden zijn in een samengroeiing, de symfyse. De symfyse is direct onder de voortanden vlak tot licht hol gekromd. Meer naar beneden wordt het profiel licht verticaal bol gekromd. Het vlak is vrij verticaal onder een hoek van 10°. De symfyse maakt een hoek van vijfenvijftig graden met de tandenrij. De achterzijde van de symfyse heeft twee versterkende beenringen, de torus superior en de torus inferior. Daartussen is een ondiepe groeve voor de aanhechting van de musculus genioglossus en de musculus geniohyoideus. Bij Mandibula III is de groeve dieper loopt de onderste ring door tot onder de eerste kies en vormt een robuuste opstaande richel in plaats van het plateau dat typisch is voor huidige apen. De wortels van de onderste hoektanden zijn aan de buitenzijde zichtbaar als verheven verticale richels, de juga, maar dat is minder bij het, onder andere daarom zo onderkende, vrouwelijke exemplaar. Onder de onderste ring ligt een opvallende verruwing voor de aanhechting van de musculus digastricus.
De zijwand van de onderkaak vormt een lichte uitholling onder de premolaren maar wordt bol onder de kiezen. Onder de vierde premolaar bevindt zich een enkel foramen mentale, een opening op een derde van de hoogte. Onder het midden of de achterzijde van de tweede kies begint een zwelling als een ondersteuning voor de ramus, de opgaande tak richting kaakgewricht. De zwelling is aanzienlijk wat erop duidt dat de tak groot was, hetgeen past bij het robuuste corpus. Bij Mandibula III vormt zich een echte richel, begrensd door een groeve voor de musculus masseter. Er is een zeer brede brede groeve voor de musculus buccinator, met tweemaal de breedte van de derde kies. Gezien de kiezen is het waarschijnlijk dat Gigantopithecus zware kauwspieren bezat. Die kunnen op de schedel verankerd zijn geweest aan een hoge schedelkam. De ramus begint echter achter de derde kies en er is een kleine tussenruimte tussen de opgaande tak en de tandenrij. De binnenzijde van het corpus is verticaal bol. Er is geen richel zichtbaar als afgrenzing van de musculus mylohyoideus.
Het gedeelte van de onderkaak onder de voortanden is zeer smal, bij Mandibula I gelijk aan vierenzeventig procent van de gezamenlijke horizontale breedte van de eerste en tweede kies en bij Mandibula II zesentachtig procent. De voortanden staan verticaal in de symfyse, in een geleidelijke boog, zonder elkaar te verdringen ofwel crowding. De tandwallen voor de kiezen buigen geleidelijk naar buiten en vooraan onder een hoek van vijftien graden weer naar binnen. De afstand tussen de gepaarde tandwallen is het grootst ter hoogte van de tweede kiezen.
Gebit bewerken

Ondergebit bewerken
De smalle lage onderste voortanden staan verticaal en zijn duidelijk kleiner dan de kiezen. Hun relatieve grootte ten opzichte van de eerste kiezen is bij de eerste voortanden kleiner dan bij alle bekende moderne basale mensapen of mensapen uit het Mioceen. Aan hun tongzijde bevindt zich een V-vormig cingulum, verdikking aan de basis, maar verder ontbreken rimpels of verticale richels. De onderste tweede voortand is breder dan de eerste. De onderste voortanden hebben extreem lange wortels, tweemaal langer dan de kronen. Hun spitsen zijn afgesleten en het slijtvlak zet zich voort op de tongzijde, soms tot de wortel toe. Dit suggereert dat de bovenste tanden zich soms aan de achterzijde van de onderste tanden bewogen.
De hoektanden zijn matig groot. De onderste hoektanden zijn laag en robuust. Hun spits verheft zich maar weinig boven het niveau van de kiezen. Hun kroon is wat afgeplat van de tongzijde naar de lipzijde. De kammen aan de kant van de tweede voortand of derde premolaar zijn slecht ontwikkeld. Daarbij moet bedacht worden dat mensapen geen eerste en tweede premolaren meer hebben — de voor zoogdieren normale eerste premolaren gingen al bij basale Anthropoidea verloren, de tweede premolaren bij basale Catarrhini — en de hoektanden dus direct aan de derde premolaren grenzen. Aan de tongzijde van de onderste hoektand bevindt zich een cingulum. De verzameling gevonden onderste hoektanden laat een bimodale verdeling zien, dus met twee grootteklassen, wat kan wijzen op een verschil tussen vrouwen en mannen. Ook deze hoektanden hebben een wortel die tweemaal langer is dan de kroon. Diastemata aan beide zijden zijn klein. Dit weerspiegelt het nauw aaneensluiten van de voortanden maar ook het weinig ontwikkeld zijn van een Canine Honing Complex, het steeds aanscherpen van de bovenste hoektand tegen de onderste derde premolaar. Gevonden bovenste hoektanden laten echter zien dat die toch de onderste tweede voortand en derde premolaar met de zijden raakten. Slijtage op de onderste hoektand begint bij het spits en zet zich voort over de zijde richting derde premolaar.
De onderste derde en vierde premolaar verschillen licht, zijn heteromorf, doordat de vierde premolaar in bouw een overgang vormt naar de kiezen. Beide hebben twee wortels. De onderste derde premolaar verschilt van de bouw die aanwezig is bij andere basale mensapen door het bezit van twee hoofdknobbels en een afwijkende indeling van het kauwvlak. De kroon is kort in de richting van de tandenrij gemeten en hoog. Aan de mesiale lipzijde is er een hoog en steil slijtvlak voor het contact met de bovenste hoektand. De dwarsdoorsnede is eivormig tot trapeziumvormig. Er zijn twee goed ontwikkelde knobbels die min of meer overdwars staan. De protoconide is iets groter en meer verheven dan de metaconide en meer richting hoektand geplaatst. De knobbels zijn gescheiden door een lengtegroeve in het midden. De groeve verbindt een putvormige uitholling aan de mesiale zijde met een breed en goed ontwikkeld basin aan de distale zijde. De verbindende kammen zijn afgerond maar goed ontwikkeld. Ze raken de opstaande distale rand van het kauwvlak alwaar zich kleine knobbeltjes ontwikkelen. De onderste vierde premolaar is 'gemolariseerd' en relatief groot. De kroon is hoog met twee hoofdknobbels en een paar knobbeltjes aan de zijde van de eerste kies. De dwarsdoorsnede is vrijwel vierkant, iets breder dan lang en wat distaal taps toelopend. De protoconide is wat omvangrijker en hoger dan de metaconide. Deze hoofdknobbels staan wijd uit elkaar en zijn verbonden door een goed ontwikkelde kam die tussen de lipzijde en tongzijde loopt. De mesiale uitholling, dus aan de zijde van de derde premolaar, is breed en vrij ondiep, iets gerimpeld, en wordt mesiaal begrensd door een opstaande tandrand. Het basin van het talonide beslaat het derde distale deel van het kauwvlak. Het is ook gerimpeld en breder maar minder verheven. Ook deze uithollingen zijn weer verbonden door een middengroeve. De distale tandranden aan de lipzijde en tongzijde zijn goed ontwikkeld en eindigen in kleine lage afgeronde knobbeltjes aan de rand tegenover de eerste kies. Er is een zwak cingulum aan de lipzijde.
.JPG)
De onderste kiezen zijn lang en tamelijk smal, ingesnoerd en aan het distale uiteinde taps vernauwd. Ze hebben hoge kronen, lage gezwollen knobbels met lage afgeronde verbindende kammen en nauwe groeven ertussen. De metaconide en protoconide zijn groot en hebben ongeveer dezelfde omvang. Een diepe groeve eindigt in een mesiale uitholling, begrensd door een hoge tandrand bezet met kleine knobbeltjes. Diepe kruisende groeven kerven de tandranden aan de tongzijde en lipzijde in. De groeve overdwars beklemtoont de insnoering van de tandkroon. De hypoconide knobbel is kleiner dan de protoconide. Aan de distale rand bevindt zich nog een klein hypoconulide, iets richting lipzijde geplaatst en begrensd door diepe groeven. Soms bevindt zich tussen dit knobbeltje en de entoconide nog een zesde hoofdknobbel, een tuberculum sextum. Dit maakt deel uit van een 'trend' bij primaten waarin knobbels aan de mesiale zijde verdwijnen en aan de distale zijde nieuw opduiken. De entoconide knobbel is kleiner dan de metaconide maar even groot als de hypoconide. Tussen entoconide en metaconide bevindt zich een extra knobbeltje. Een kort kammetje tussen entoconide en hypoconide begrenst een putvormige uitholling aan de distale zijde. De distale tandrand is laag en slecht ontwikkeld, voorzien van kleine knobbeltjes. Het basin van de talonide sector, ongeveer in het midden van het kauwvlak, is lang, laag, ondiep, smal en gerimpeld wat door het dikke glazuur weer nieuwe knobbeltjes oplevert. Daarbij loopt het basin mesiaal uit in een dikke zwelling die ook weer een knobbelvormige structuur oplevert. Al met al is het patroon er een met vier hoofdknobbels aan de randen met in de richting van de tandenrij ertussen een rij van kleinere knobbels. Bij afgesleten kiezen blijft daar echter soms weinig van over en zijn er grote slijtvlakken waar het dentine zichtbaar is. Een cingulum aan de lipzijde ontbreekt of is slechts gering aanwezig.

De eerste kies is typisch kleiner dan de tweede, met gemiddeld 86,1 procent van de oppervlakte van het kauwvlak. De derde kies is meer variabel maar vaak kleiner dan de tweede, gemiddeld met ongeveer negentig procent van het kauwvlak. De wortels van de onderste kiezen zijn opmerkelijk lang. Ze hebben een absolute lengte welke die bij moderne mensapen verre overtreft.
Bovengebit bewerken
Het bovengebit is alleen bekend van losse tanden. Omdat er daar zoveel van gevonden zijn, bezitten we exemplaren voor alle tandposities. De bovenste voortanden lijken op de onderste en zijn dus relatief klein, kleiner dan bij moderne basale mensapen. De geringe breedte van hun tandenrij doet vermoeden dat het foramen incisivum in het verhemelte centraal ligt en enkelvoudig is als bij orang-oetans en de moderne mens, afwijkend van de dubbele openingen, de fenestra palatinae, bij chimpansees en gorilla's. De eerste voortand heeft een vierkante dwarsdoorsnede. De tweede voortand heeft een goed ontwikkeld cinguluem aan de tongzijde. De tweede voortand is extra klein en heeft twee derden van de lengte van de eerste voortand. Dat lijkt op de toestand bij orang-oetans.
De bovenste hoektanden zijn laag maar robuust en vermoedelijk langer dan de onderste hoektanden. Ze zouden beneden de rij van de kiezen hebben uitgestoken. Ze werden kennelijk aangescherpt tegen de onderste derde premolaren. Terwijl de zijde aan de kant van de twee voortand is afgerond, vormt de distale zijde een scherpe kiel naast een ondiepe verticale groeve. Er is nog een spoor van een cingulum aan de tongzijde. Ook deze hoektanden zijn te verdelen in twee grootteklassen, met vermoedelijk de grootste bij de mannen.
.JPG)
De bovenste premolaren zijn breed met een hoge kroon. De derde premolaar is iets groter dan de vierde. Hij heeft een driehoekige dwarsdoorsnede, breder aan de lipzijde. De vierde premolaar heeft een eironde dwarsdoorsnede. Beide premolaren hebben twee hoofdknobbels waarbij de paracoon veel groter is dan de protocoon. De knobbels worden verbonden door een of twee lage afgeronde kammen. Die begrenzen een klein centraal basin en spleetvormige foveae aan de mesiale en distale zijde. Deze uithollingen worden verbonden door een onregelmatige insnijding. Rimpels in het email vormen knobbeltjes in de centrale en distale uithollingen. De derde premolaar heeft een matig ontwikkeld cingulum aan de tongzijde; bij de vierde premolaar is dit vrijwel afwezig. Beide premolaren hebben drie wortels. De grootste daarvan bevindt zich aan de lipzijde; twee kleinere worteltjes staan aan de tongzijde.
De bovenste kiezen hebben een rechthoekig tot ruitvormig profiel van het kauwvlak. Ze zijn maar iets breder dan lang en lopen distaal taps toe. Hun insnoering is minder dan bij de onderste kiezen. De kronen zijn hoog en blokvormig met steile wanden maar matig hoge knobbels. De knobbels zijn groot, afgerond en gezwollen, veel van het kauwvlak opvullend en ook hier gescheiden door diepe groeven. Protocoon en paracoon zijn beide even groot maar veel groter dan de andere hoofdknobbels. De metacoon is kleiner maar goed ontwikkeld, gelegen op de hoek tussen de lipzijde en de distale kant. De crista obliqua is hier een weinig opvallende zwelling, schuin doorsneden door een groeve. Deze kam loopt mesiaal uit in de hypoprotocrista, in de vorm van een knobbeltje op het midden van het kauwvlak. De hypocoon is de kleinste hoofdknobbel maar nog steeds aanzienlijk van omvang. De kammen zijn zwak ontwikkeld en afgerond. De middengroeve is diep ingekerfd, omgeven door de knobbeltjes van de kammen. Een inkerving daar doorheen verbindt de middengroeve met het talonide basin aan de distale zijde. Evenzo is de fovea aan de distale zijde met dit basin door een groeve verbonden. Door rimpels in het glazuur vormen knobbeltjes aan de rand van de groeven en het basin. Een cingulum aan de tongzijde is afwezig of heeft de vorm van een zwak spoor. Ook hier is de eerste kies kleiner dan de tweede en is de derde kies variabel maar kleiner. De derde kies is korter in de richting van de tandenrij met een bolle distale rand en een sterk gereduceerd metacoon en hypocoon.
Grootte en glazuur van de kiezen bewerken

De kiezen zijn zeer groot, de grootste die van mensapen bekend zijn. Vergeleken met Gorilla gorilla, de grootste huidige mensaap, is de oppervlakte van het kauwvlak van de bovenste kiezen 57,8 procent hoger en dat van de onderste drieëndertig procent. Alleen Indopithecus en Ouranopithecus turkae benaderen deze grootte. De kiezen zijn ook meer hypsodont, met een grotere relatieve kroonhoogte, dan bij alle andere bekende mensapen. De kiezen hebben daarbij in absolute zin het dikste tandemail van enige bekende mensaap, maximaal zes millimeter en gemiddeld 2,5 tot 2,9 millimeter. Relatief tot de tand als geheel is het echter dunner dan bij Graecopithecus freybergi, Lufengpithecus lufengensis, Australopithecus africanus en Paranthropus robustus. Bij de bovenste kiezen is het email het dikst op de knobbels aan de tongzijde, bij de onderste kiezen aan de lipzijde. Daar is de slijtage het grootst. Het email is overigens vrij gelijkmatig verdeeld, meer dan bij moderne mensen. De grenslaag met het dentine heeft een vrij gelijkmatig reliëf, net als bij orang-oetans. Het email bestaat aan de buitenkant uit Patroon-I-prismata en intern uit Patroon IIIa-prismata.
Postcrania bewerken
Van de postcrania, de delen 'achter de schedel', is niets teruggevonden. Dat maakt het lastig romp en ledematen te reconstrueren. De vorm is slechts af te leiden uit de fylogenetische positie, in de evolutionaire stamboom, en de grootte van het hoofd. De schedel was duidelijk zwaar en moet gedragen zijn door een forse nek. De vorm van de verdere ruggengraat is onzeker. Moderne chimpansees en gorilla's hebben een verkorte onderrug met een verminderd aantal lendenwervels. Dit vergroot de stabilisatie bij het klimmen. Gigantopithecus was aan deze vormen niet nauw verwant en indien hij zeer robuust van bouw en voornamelijk een grondbewoner was, zal hij een dergelijke specialisatie niet nodig gehad hebben. De onderrug kan dus lang en soepel geweest zijn met vijf lendenwervels en vijf sacrale wervels. Een soepele rug zou een overeenkomst geweest zijn met orang-oetans, de vermoede nauwste nog levende verwanten. Die hebben zelf ook aanpassingen aan het klimmen, zoals zeer lange armen en het typerende 'dubbele heupgewricht'. Ook hiervoor geldt dat die overbodig waren voor een zware grondbewoner.
Fylogenie en afstammingsgeschiedenis bewerken

Von Koenigswald stelde in de oorspronkelijke beschrijving van 1935 dat Gigantopithecus nauw verwant was aan Sivapithecus uit het middelste Mioceen van India. In 1939 was Robert Broom de eerste die een band opperde met mensachtigen, australopitheken, op grond van de gelijkende grote kiezen met dik glazuur, met name Plesianthropus en Paranthropus. Die hypothese herhaalde hij in 1946. Franz Weidenreich sloot zich in 1937 bij de interpretatie van Von Koenigswald aan. Toen die laatste door de Japanners geïnterneerd was, kon Weidenreich zijn studie van het taxon voortzetten in New York, bij het American Museum of Natural History. Daarbij betrok hij ook de extra zes tanden die gevonden waren. In 1945 ging hij over tot het kamp Broom en beschouwde Gigantopithecus als een lid van de Hominidae in de enge zin, dus de tegenwoordige Hominini. Het zou aldus een mensachtige geweest zijn die hij zag als een plausibele voorouder van Homo erectus. Het wezen zou er dan uitgezien hebben als een tweevoetige reus. Von Koenigswald was het in 1952 eens met de status van mensachtige maar wierp tegen dat het dier te gespecialiseerd was om een voorloper van de mens geweest te zijn. Moderne mensen zouden dus geen reuzenfase in hun ontwikkeling hebben gehad, een romantische gedachte die velen aansprak. Von Koenigswald hield echter vast aan een connectie met India, Indopithecus als een oudere verwant van Gigantopithecus ziend.
Tot in de jaren tachtig zouden sommige westerse onderzoekers de soort als een mensachtige zien. De Chinese onderzoekers daarentegen bleven Gigantopithecus als een basale mensaap beschouwen. Dat kreeg in het westen meer aanhang na de benoeming van Gigantopithecus bilaspurensis. Toch sloot voor velen de verwantschap met Indiase en Pakistaanse vormen het behoren tot de menselijke tak niet uit. Het was namelijk populair Ramapithecus als de voorloper van de mens te beschouwen. Pas toen duidelijk werd dat die nauw aan de orang-oetans verwant was en waarschijnlijk identiek aan Sivapithecus, verhuisde Gigantopithecus naar de Ponginae. Een minderheidspositie nam Jeffrey Schwartz in die in 1987 met zijn The Red Ape stelde dat de orang-oetan nauwer aan de mens verwant was dan de chimpansee of gorilla. Dit viel echter niet te verenigen met de resultaten van het onderzoek naar eiwitten en DNA.
De huidige consensus is dat Gigantopithecus nauwer verwant is aan de orang-oetan en het eveneens uitgestorven maar veel kleinere geslacht Sivapithecus. De orang-oetan is dan het naaste nog levende familielid. In 2019 werd dit bevestigd door onderzoek naar fossiele eiwitten. Experts op het gebied van paleoproteomics in Denemarken hebben monsters van tandglazuur verkregen van een bijna 1,9 miljoen jaar oude kies uit China en reconstrueerden verschillende eiwitten van Gigantopihecus blacki. Het proteoom was met een factor vijf het oudste dat tot dan toe uit een fossiel werd gewonnen. De reconstructies werden op basis van de gegevens in eiwitdatabases vergeleken met daar geregistreerde eiwitten van andere apen. Dit bevestigde dat Gigantopithecus een zustertaxon van de orang-oetan (geslacht Pongo) is en dat de laatste gemeenschappelijke voorouder van Gigantopithecus en orang-oetan twaalf tot tien miljoen jaar geleden leefde.
Zhang wees er in 2017 op dat de band met de orang-oetan onzeker was. Gigantopithecus deelt sommige synapomorfieën met Pongo zoals kiezen met dik glazuur, een hoog reliëf van de grenslaag met het dentine, ondiepe uithollingen op het kauwvlak en lage knobbels. Het probleem is dat dit alleen apomorfieën zijn als vergeleken wordt met moderne soorten. Veel basale mensapen uit het Mioceen en aapmensen uit het Pleistoceen hebben dezelfde kenmerken. De combinatie kan zelfs de gemeenschappelijke voorouder van alle Hominidae getypeerd hebben en dus een symplesiomorfie zijn die geen informatie biedt over de verwantschappen. Alles hangt dus af van de voorgestelde overeenkomsten met Sivapithecus. Kleine voortanden, kiesvormige premolaren en grote kiezen komen echter opnieuw bij veel mensapen voor en leveren geen eenduidig bewijs op. De positie van Gigantopithecus berust dus eigenlijk voornamelijk op algemene scenario's over de hominide evolutie, waarin het om geografische redenen zoals het ontbreken van Hominini in Oost-Azië die ouder zijn dan twee miljoen jaar het meest plausibel is dat het om een pongine gaat. Slechts de vondst van meer volledige skeletten van Gigantopithecus, of van directe voorlopers, kan uitsluitsel bieden.
Het volgende kladogram toont de positie van Gigantopithecus in de evolutionaire stamboom volgens een studie van Begun uit 2007.
| Hominoidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Levenswijze bewerken
Leefgebied bewerken

De vindplaatsen van Gigantopithecus liggen voornamelijk in gebieden die tegenwoordig een subtropisch klimaat hebben. Hainan ligt in een tropisch klimaat. Ondanks de klimaatveranderingen tijdens het Pleistoceen is het waarschijnlijk dat het lokale klimaat indertijd niet sterk van het huidige afweek. Dit wordt bevestigd door de in de grotten gevonden fossiele fauna. Die uit de zuidelijke grotten is typisch subtropisch terwijl in de meest noordelijke grotten meer koudebestendige soorten te vinden zijn, typisch voor het palearctisch gebied. Onder deze fauna bevinden zich veel zoogdieren die bosbewoners zijn. Daar maken veel smalneusapen deel van uit zoals Macaca, Rhinopithecus, Pygathrix, Trachypithecus, Nomascus en Pongo, dus orang-oetans. Er was dus een rijke gemeenschap van klimmende apen. Een soortgelijke combinatie is in moderne bossen niet meer te vinden; de nauwste benadering vormen de regenwouden van Noord-Vietnam. Misschien dat Gigantopithecus de niche vulde van bodembewonende aap. Fossiele boombladeren wijzen op een warm-gematigd tot subtropisch klimaat en een begroeiing door loofbomen waarvan sommige bladverliezend. Het bos werd gedomineerd door berken, eiken en chinkapin met een ondergroei van struiken en kruiden waaronder varens, grassen en tweezaadlobbigen. Onderzoek naar de koolstof- en zuurstofisotopen in het tandglazuur wees erop dat de habitat subtropisch was, vochtig en dicht bebost met een gesloten bladerdek. Een moderne analoog zou het Dinghushan Biosphere Reserve zijn, een natuurreservaat in China met een gemiddelde jaarlijkse temperatuur van eenentwintig graden Celsius (dat wijst dus op een veel hogere gemiddelde maximale dagtemperatuur) en door de moesson een gemiddelde jaarlijkse regenval van 1900 millimeter.
Een schaduwrijk bos kan hebben geleid tot een geringere pigmentatie van de huid en een vacht die lichter in kleur was. Vaak wordt Giganthopithecus met rood haar afgebeeld, net als bij orang-oetans. Die hebben typisch een erg dichte of lange vacht die bij mannen deel uitmaakt van een heel arsenaal aan intimiderende pronkorganen zoals snorren, baarden, wangkussens en keelzakken. Het is mogelijk dat, maar zeer onzeker of, ook Gigantopithecus hiermee uitgerust was. De hoge onderkaak, extra hoog bij mannen, zou een aanwijzing kunnen zijn voor een keelzak, met toegang van de keelholte uit, die bij basale mensapen vaak sterk ontwikkeld is. Bij de moderne mens is die verdwenen. Een lange vacht kan bij een groot dier in een warm klimaat het gevaar van oververhitting met zich meebrengen. Dezelfde lichaamsomvang echter kan ook als een buffer gediend hebben tegen een gevaarlijke abrupte temperatuurstijging.
Dieet bewerken
.JPG)
De onderkaak van Gigantopithecus, robuust, hoog en naar achteren verdiepend, met een verstevigde symfyse, was in staat een hoge belasting te weerstaan bij het kauwen van taai of hard voedsel. Dergelijke aanpassingen zijn echter niet eenduidig. Een hoog corpus is aangetroffen bij primaten die bladeters, zaadeters en krakers van harde noten zijn. Een hoge achterste kaak komt voor bij bladeters maar ook zaadeters. De hoge tanden met dik email dat daarbij soms flink versleten is, wijzen op voedsel dat een forse wrijving opwekt. De kiesvormige premolaren, de enorme kiezen en de zeer lange en robuuste tandwortels duiden op een zwaar kauwproces bij het verwerken van volumineuze en vezelrijke kost. De lage knobbels, verbonden door zwakke kammen, waren niet geschikt om bladeren, inclusief grassen, te verwerken. De tanden tonen een hoge mate van tandbederf — elf procent van de kiezen heeft gaatjes, vergelijkbaar met dat bij moderne chimpansees, wat veroorzaakt kan zijn door een suikerrijk dieet, bijvoorbeeld van vruchten. De voortanden zijn echter klein, terwijl die bij fruiteters typisch groot zijn. Hun geringe lengte duidt erop dat het voedsel geen voorwerking nodig had om gekauwd te worden. Daartegen spreekt weer het feit dat ze meestal flink versleten zijn.
Gigantopithecus leefde vermoedelijk mede in bamboebossen. Sinds de jaren zeventig opperden sommige onderzoekers, waaronder Tim White, dat hij zich voedde met bamboebladeren als de huidige reuzenpanda, Ailuropoda melanoleuca. Overeenkomsten zijn de robuuste kaken, de bunodonte, dus afgerond blokvormige, tanden en de vele knobbels op het kauwvlak. Een probleem is dat de reuzenpanda gezien fossiele vondsten al tijdens het Pleistoceen in het gebied aanwezig was en beide soorten elkaar dus beconcurreerd zouden hebben. Daarbij wijzen de hogere tanden en het dikkere glazuur bij Gigantopithecus op andere aanpassingen en dus misschien ook op een ander dieet. Een andere analogie is gevonden in de berggorilla die ook vezelrijk materiaal eet.
Een manier om het dieet meer direct vast te stellen is het onderzoeken van het glazuur op minuscule harde plantendelen, fytolieten. In 1990 bleek het Ciochon dat er zich in het glazuur zeldzame resten van grassen bevinden maar talrijke fytolieten van de moerbeifamilie. Dergelijke Moraceae omvatten niet alleen moerbeien maar ook de broodvrucht, de doerian en vijgen. Het gaat om struiken met veel voedzame vruchten, ideaal voor een grote bodembewoner. In 2014 werden deeltjes zetmeel gemeld die zouden wijzen op een dieet van wortels of knollen. Ook het patroon van afzetting van tandplak wijst daarop.
Een andere methode is het met een microscoop waarnemen van kleine krasjes en putjes in het glazuur, de micro wear. De putjes komen volgens een studie uit 1994 overeen met die op tanden van chimpansees en waren veel ondieper dan die bij orang-oetans die veel hardere objecten kraken. De krasjes leken niet op die bij typische bamboe-eters. In 2014 bleek dat exclusief C3-planten werden gegeten. Zhang concludeerde dat Gigantopithecus een generalist was die verschillende typen voedsel at uit meerdere plantenlagen: wortels en paddenstoelen van en uit de bodem, struiken en uit het bladerdak naar beneden gevallen vruchten. Dat laat de vraag onverlet waarom de kiezen zo groot moesten zijn. Bij robuuste australopitheken, die ons voor dezelfde problematiek stellen omdat ze grote kiezen hebben maar het glazuur op de dikte na niet bijzonder afwijkend is, wordt dit verklaard door of aan te nemen dat de enorme molaren het innemen van voedsel van grote kwantiteit mogelijk maakten in plaats van een speciale kwaliteit, of door te veronderstellen dat ze toestonden in tijden van droogte terug te vallen op knollen. Dat laatste lijkt in een subtropisch regenwoud niet relevant te zijn geweest.
Een studie van de koolstofisotopen in het tandglazuur, in 2017 gepubliceerd door Hervé Bocherens, concludeerde dat Gigantopithecus net als de orang-oetan bladeren at maar vooral vruchten en, in tegenstelling tot een pandabeer, niet gespecialiseerd was in bamboe. Het uitsterven van Gigantopithecus zou plaatsgevonden hebben toen de bossen verdwenen tijdens de ijstijdfasen van het Pleistoceen en werden vervangen door meer open savannes, zodat de voedselvoorziening verslechterde. Terwijl de voorouders van mensen die in Afrika woonden tegelijkertijd grassen en wortels als voedsel gebruikten, die ook overvloedig aanwezig waren in de Aziatische savannes, paste Gigantopithecus zich blijkbaar niet snel genoeg aan de veranderende omgeving aan. De orang-oetan had het voordeel dat dit dier zijn stofwisseling kan verlagen om perioden met voedselschaarste te overleven. Het uitsterven zou wellicht pas honderdduizend jaar geleden voltooid zijn.
Pei dacht in 1957 dat Gigantopithecus een jager was. Fossielen van hoefdieren zouden slechts door de grote aap in de slecht toegankelijke grotten gesleept kunnen zijn. Deze hypothese is nu geheel verlaten. Het is echter waarschijnlijk dat het dieet zo ver als mogelijk door het eten van karkassen en ongewervelden met dierlijke eiwitten werd aangevuld.
Evolutionaire verandering bewerken
Het grote aantal tanden die gekoppeld konden worden aan precieze grotten waarvan de datering bepaald kon worden, stelde Zhang in staat een statistische analyse uit te voeren van evolutionaire veranderingen gedurende het bestaan van Gigantopithecus. Hij verdeelde ze in twee groepen, uit het vroege Pleistoceen en uit het middelste Pleistoceen. Hij concludeerde in 1982 en 1983 dat de tanden in de loop der tijd aanzienlijk groter waren geworden. In 2015 was het aantal kiezen gegroeid tot 1218 en kon de analyse herhaald worden. Alle premolaren en kiezen in de tandenrij werden groter. Daarbij werden de eerdere conclusies bevestigd. De toeneming in grootte kan een groei in de lichaamsomvang weerspiegelen. Er zijn bekende factoren in een selectiedruk om groter te worden. Grotere dieren hebben een voorsprong op soortgenoten bij het zoeken van voedsel of een partner en zijn beter beschermd tegen roofdieren. Een langer darmstelsel kan beter voedsel verteren. Een grotere massa maakt een dier ook minder gevoelig voor voedseltekorten en temperatuurswisselingen.
De veranderingen kunnen er echter ook op duiden dat de kiezen groter werden ten opzichte van de kaak als geheel. Dat kan duiden op een verandering in het dieet. Wellicht werden de bossen geleidelijk vervangen door graslanden en waren kiezen met dikker glazuur nodig om grassen te eten. Het kan ook zijn dat Gigantopithecus is uitgestorven omdat hij zich daaraan niet snel genoeg kon aanpassen. Grote dieren hebben kleinere populaties en planten zich langzamer voort wat ze meer uitstervingsgevoelig maakt.
Behalve in grootte, veranderden de tanden ook in vorm. De tanden uit de grot van Heijang, die 400 000 tot 320 000 jaar oud zijn, bleken een grenslaag tussen het dentine en het glazuur te hebben die meer reliëf toont. De kammen op het kauwvlak zijn scherper en beter gevormd. Dat geldt vooral voor de bijkomende kammen die van de paracoon van de premolaren uitstralen en welke ook talrijker zijn. Anders dan bij oudere tanden, is bij de derde premolaren geen verbinding meer tussen de voorste tandrand van de preprotocrista en de kam tussen de parastyle en de paracoon. Bij de vierde premolaren ontbreekt de verbinding met de preparacrista zodat er een trog ontstaat die de mesiale tandrand onderbreekt. Al met al neemt de complexiteit van het kauwvlak toe wat wellicht een aanpassing is voor het scheuren van grassen.
Voortbeweging bewerken
Door zijn grootte was Gigantopithecus vermoedelijk niet in staat om veilig hoog in de bomen te klimmen. Takken dreigden af te breken en een val van enige meters zou hij niet overleefd hebben. Het lijkt niet aannemelijk dat hij aan takken slingerde. In zijn aan de bodem gebonden levensstijl geleek Gigantopithecus veel op de gorilla. Dat roept weer de vraag op hoe hij zich op de grond voortbewoog. Gorilla's en chimpansees tonen een specialisatie die niet van andere mensapen bekend is: het knuckle walking. Bij oppervlakkige beschouwing lijkt het alsof ze op hun vuisten lopen. Dat zou echter betekenen dat de vingers in hun geheel naar achteren gebogen worden. In feite worden de eerste kootjes van de vingers gestrekt zodat ze vooruit steken. De sterk verlengde tweede kootjes maken daar een hoek van 180° mee en steken naar achteren. De menselijke hand is daartoe niet in staat. Bij chimpansees en gorilla's hebben de onderste uiteinden van de eerste kootjes een speciale buiging naar achteren om die hoek mogelijk te maken. Het gewicht rust dan op de tweede kootjes die voorzien zijn van dikke dempende weefselkussens en eeltlagen. De extreme specialisatie van dit systeem is een argument om het niet bij Gigantopithecus aanwezig te achten. Daartegen spreekt echter dat het bij chimpansees en gorilla's apart schijnt te zijn geëvolueerd omdat hun gemeenschappelijke voorouder het kennelijk niet bezat gezien het feit dat er bij mensachtigen geen spoor van is aangetroffen. Orang-oetans lopen vaak met de vingers naar achteren geklapt ten opzichte van de middenhand en misschien dat Gigantopithecus dat ook deed. Een laatste mogelijkheid is dat hij nog de oorspronkelijke loopwijze van de apen bezat, met de vingers plat naar voren op de grond gehouden.
Gigantopithecus hoeft niet snel te zijn geweest. Hij had als voordeel dat vermoedelijk geen enkel niet-menselijk roofdier groot genoeg was om volwassen exemplaren te kunnen doden. Kinderen waren denkelijk wel in staat te klimmen. Gigantopithecus leefde echter op het eind van zijn bestaan gelijktijdig met Homo erectus. Deze mensensoort was ook niet snel maar had zich gespecialiseerd in uithoudingvermogen: hij kon in een gestage achtervolgingsjacht zijn slachtoffers uitputten. Mensen kunnen met hun lange benen en vele zweetklieren in een naakte huid, uren achter elkaar hollen maar Gigantopithecus zal na een kwartiertje op apegapen hebben gelegen, zeker omdat hij zo zwaar was en in een heet vochtig klimaat leefde. Misschien dat hij tot een zekere mate van primitief werktuiggebruik in staat was en zich wat kon verweren met stokken en stenen maar een solitaire man zal het niet lang hebben uitgehouden tegen een met speren bewapende jachtgroep van Homo erectus.
Seksuele dimorfie bewerken

De gevonden onderkaken tonen een hoge mate van de seksuele dimorfie, verschillen tussen de seksen. Bij Mandibula III zijn de hoektanden veel langer en hebben een grotere dwarsdoorsnede dan bij Mandibula I. Dat is verklaard door aan te nemen dat de eerste van een man is en de tweede van een vrouw. Bij mensapen hebben de mannen typisch de grotere hoektanden. Een zelfde bimodale verdeling is vastgesteld bij de losse bovenste hoektanden die gevonden zijn. Bij een partij is een gemiddelde lengte van 21,1 millimeter gemeten bij mannen en 15,4 millimeter voor vrouwen. De mannelijke hoektanden zijn dus zevenendertig procent langer wat een hogere verhouding is dan bij alle andere bekende Hominoidea, behalve de gorilla's.
De schedel van mannen lijkt ook in het algemeen groter te zijn geweest. Ter hoogte van de eerste kies heeft Mandibula III een 40% grotere hoogte dan Mandibula III. Dat is meer dan bij alle bekende moderne Hominidae, inclusief de gorilla's waar de hoogte maar een vijfde tot zesde meer bedraagt. Hier kan uit worden afgeleid dat mannen ook veel zwaarder waren. Dat duidt op een sterke competitie tussen de mannen onderling. Het is echter de vraag of die met de hoektanden werd uitgevochten want die waren maar klein ten opzichte van de kaken als geheel. Al te dreigend kan het ontbloten van de hoektanden dus niet zijn overgekomen. Bij andere soorten is zo'n toestand wel gezien als een teken voor een laag agressieniveau. Wellicht was er een grote mate van seksuele selectie door de vrouwen, met een sterke voorkeur voor zware mannen.
Uit seksuele dimorfie worden ook wel conclusies getrokken over de sociale structuur bij primaten. Grote mannen kunnen erop wijzen dat men in een harem leefde, met één man en meerdere vrouwen, zoals bij gorilla's. Ze passen minder bij een leven in gemengde groepen als bij chimpansees. Een ander model verschaffen de gibbons die in paren leven. De vermoedelijk nauwste verwanten, de orang-oetans, zijn onder primaten zeer uitzonderlijk omdat ze niet sociaal zijn maar solitair leven. Ook dat is een plausibel scenario voor Gigantopithecus: een grote man kan een ruim territorium verdedigd hebben dat voldoende voedsel verschafte voor zijn grote lichaam.
Groei bewerken
De groei moet een langere groeifase gekend hebben dan bij andere mensapen. In 2003 werd geschat dat het email op de kies van het blijvende gebit zeshonderd tot achthonderd dagen nodig had om afgezet te worden, met elf dagen per groeilijn. De vorming van een hele tandkroon zou 4,09 jaar gevergd hebben. Dat ligt dicht bij de bovengrens bij moderne mensapen. De studie uit 2019 naar het eiwit uit de 1,9 miljoen oude kies concludeerde dat alpha-2-HS-glycoproteine een rol speelde bij de permineralisatie van het glazuur.
Kryptozoölogie bewerken

Gigantopithecus is ook een rol gaan spelen in de kryptozoölogie, de zoektocht naar soorten waarvan het bestaan nooit is aangetoond. Op verschillende plaatsen ter wereld zijn er legenden of ankedoten over reusachtige apen. Veel cryptids betreffen wezens die gezien de huidige stand van de wetenschap nauwelijks kunnen bestaan zoals een plesiosauriër in Loch Ness of een sauropode in de Congo: ze zouden het restant moeten zijn van diergroepen die al zesenzestig miljoen jaar geleden zijn uitgeroeid. Voor de verschrikkelijke sneeuwman ofwel yeti en Bigfoot geldt echter dat ze best in de huidige fauna zouden passen. Alleen hun grootte is opmerkelijk. Gigantopithecus bewijst in ieder geval dat zeer grote mensapen echt bestaan hebben en zijn moment van uitsterven, 320 000 jaar geleden, is in geologisch opzicht zeer recent. China ligt daarbij dicht bij Tibet en zou in Beringia een verbinding gehad hebben waarlangs een grote en dus relatief koudebestendige aap naar Noord-Amerika kon trekken.
De eerste welke dit opviel was in 1952 Bernard Heuvelmans. In 1960 kreeg de hypothese wat meer gewicht door de zoöloog Wladimir Tschernezky, die in Nature een foto van een vermeend yetispoor besprak genomen door de bergbeklimmers Michael Ward and Eric Shipton. Tschernezky oordeelde dat de yeti liep als een mens en leek op Gigantopithecus. Hij dacht dus dat die op een mensachtige leek. Antropoloog Grover Krantz heeft tot zijn dood in 2002 de hypothese verdedigd dat Gigantopithecus, de yeti en bigfoot identiek waren. Krantz had in 1969 het werk van Franz Weidenreich gelezen maar verwaarloosde latere literatuur. Hij dacht dat Gigantopithecus een tweevoeter was die via allerlei aapmensen in vermeende moderne rassen evolueerde die dus een zeer hoge ouderdom gehad zouden hebben, een gedachte die bij twintigste-eeuwse racisten zeer populair was. Ook de belangstelling van Weinert voor dit onderwerp had een duidelijk racistische inslag. In januari 1985 probeerde Krantz afgietsels van voetafdrukken van bigfoot naar voren te schuiven als het type van een ichnospecies Gigantopithecus blacki, een artikel voordragend bij een vergadering van de International Society of Cryptozoology. Het artikel werd echter nooit gepubliceerd omdat Krantz geen holotype kon laten zien. In 1985 opperde hij als alternatieve naam voor bigfoot de combinatie Gigantopithecus canadensis. De meeste wetenschappers zien die naam als overbodig omdat ze niet geloven dat de bigfoot echt bestaat. De waarnemingen kunnen plausibel verklaard worden als ontmoetingen met op hun achterpoten staande beren. Zelfs als Gigantopithecus pas honderdduizend jaar geleden uitstierf, is het ook niet waarschijnlijk dat een collectieve herinnering aan hem de bron is van de legenden over reuzenapen want die tijdspanne is een orde van grootte hoger dan de maximale tijd waarbinnen volgens de antropologie nog informatie overgedragen wordt via verhalen.
Populaire cultuur bewerken
In de Disneyfilm The Jungle Book wordt King Louie als een gigantopithecus opgevoerd.
Galerij bewerken
-
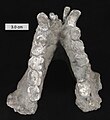 Tanden van Gigantopithecus
Tanden van Gigantopithecus -
 Tanden van Giganthopithecus blacki
Tanden van Giganthopithecus blacki
(op de achtergrond prof. Friedemann Schrenk)

| Bronnen, noten en/of referenties
Literatuur
Noten
|